Глава VI Дифференцование полов в органическом прогрессе
Выше изложен в общих чертах ход полового подбора при действии простой формы наследственности; теперь посмотрим, какие последствия произойдут от тех осложнений в процессе наследственности, которые мы затем констатировали? В половом подборе, конечно, первоклассную важность имеет ограничение наследственности полом. Под влиянием этого-то закона[27], по Дарвину, половой подбор производит вторичные половые признаки. Посмотрим же на примере, действительно ли это так? и в каких случаях это может иметь место? Итак, возобновим вычисление и будем в него вводить мало-помалу различные осложнения; для начала же предположим, что действует только закон ограниченной полом наследственности, как он изложен у Дарвина в его "Происхождении человека и половом подборе". Мы имеем, положим, 40 000 особей установившегося вида или породы; по-прежнему численное отношение обоих полов одинаково, как 100: 100; по-прежнему половина самок раньше созревает и плодовита, т. е. приносит по два детеныша, а другая, малоплодная, приносит по одному; по-прежнему молодые достигают половой зрелости в один год и расстояние половых периодов тоже год; наконец, по-прежнему естественные условия держат вид в одном количестве, не допуская размножаться. После мы введем этот ограничивающий размножение естественный подбор в расчет и рассмотрим, как он отражается и как совершенно видоизменяет эффекты полового подбора, но пока мы для простоты задачи предположим, что борьба за существование действует безразлично на все особи, не препятствуя половому подбору. Затем, конечно, мы предположим, что измененные условия существования вызвали в виде игру изменчивости, которая выразилась, между прочим, тем, что известное количество особей приобрело особую окраску, понравившуюся самкам вида; так как мы рассматриваем действие наследственности, ограниченной полом, то нам полезно ввести гипотезу, что первоначально этот новый признак появился только у одного пола, положим, у самцов. Это уклонение проявилось отчасти в молодости и детстве, отчасти в зрелом возрасте, и для простоты задачи предположим, что этих уклонений произошло поровну, а всего уклонившихся 20 % общего количества приплода, так что выходит 3000 рано уклонившихся и 3000 приобретших уклонения в зрелом возрасте. Нижеследующая таблица показывает результат этого приплода.

В последней вертикальной графе выставлено, сколько должно остаться особей разного рода к следующему половому периоду, когда борьба за существование опять приведет вид к его норме в 40 000 особей. Так как с приплодом оказалось 70 000, то выжить должны только 4/7 особей каждого сорта, сообразно с чем и сделан расчет в предположении, что естественный подбор вовсе не влияет на признак, подбираемый самками. Таков состав первого поколения, в котором появилось предположенное отклонение у самцов; вычислим теперь второе, третье и т. д. до десятого, тогда мы получим нижеследующие таблицы[28].
Таблица приплода 10-ти поколений
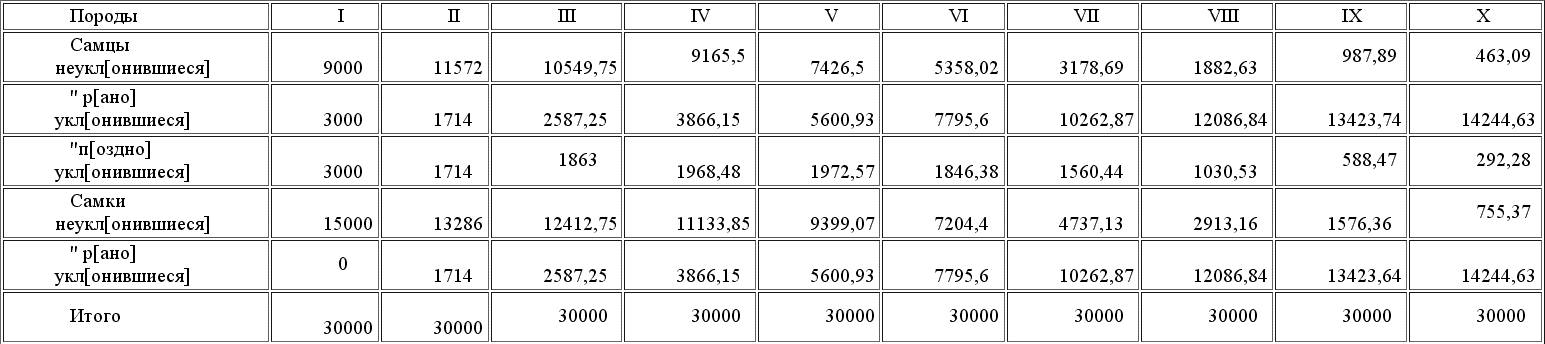
Таблица состава 10-ти поколений к периоду размножения
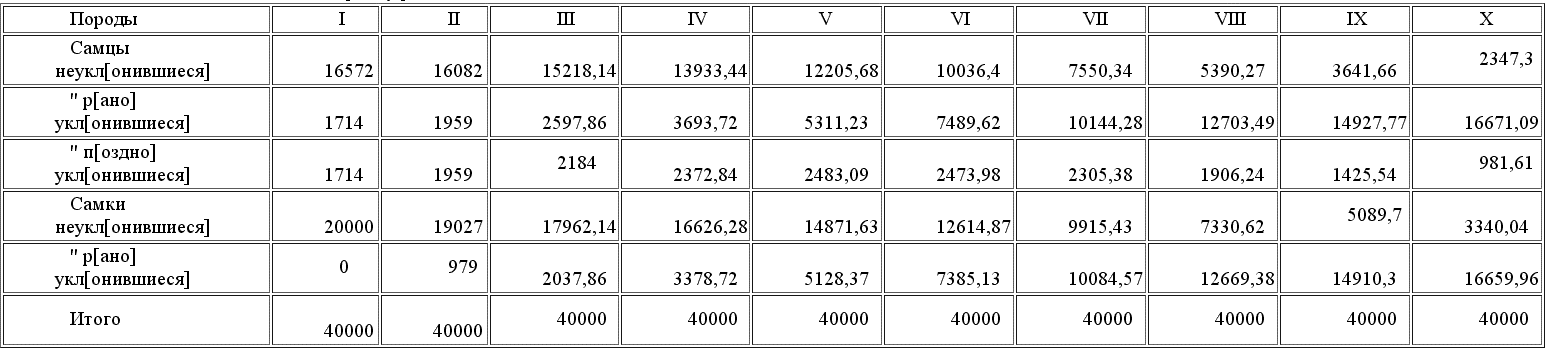
Выводы ясны; таблицы красноречивы сами по себе и многих комментариев не требуют. Очевидно, что рано или поздно процесс полового подбора должен вытеснить не только неуклонившиеся формы, не привлекательные для самок, но и поздно уклонившиеся, равно привлекательные, как и ранние уклонения. Такой результат происходит от двух причин: 1) от абсолютной неравночисленности размножающих уклонение особей, происходящей вследствие нашего молчаливого допущения, что только потомки мужеского пола поздно уклонившихся особей передают этот признак своим детям. Насколько мы имели право сделать подобное предположение и в каких пределах оно согласно с действительностью, мы разберем немного позже. 2) Вторая причина большей быстроты размножения рано уклонившихся особей заключается в том, тоже молчаливо допущенном условии, что в то время как потомки рано уклонившихся особей все наследуют родительский признак, между потомками поздно уклонившихся часть, происшедшая от спариванья с рано уклонившимися самками, увеличивают собою ряды не отцовской, а материнской породы. Основания для этих допущений мы сейчас рассмотрим, но если они правильны, то становятся вполне понятны результаты наших вычислений, что станет еще нагляднее из нижеследующих таблиц плодовитости и приращения разных пород. Общее число рождений в нашем гипотетическом виде 30.000 особей при численности всего вида в 40.000, так что на 100 особей всех пород рождается приплода 75 детенышей всех пород. Таков % рождаемости нашего вида вообще, а вот таблица рождаемости каждой породы, вычисленная для каждого поколения.
На 100 взрослых особей каждой породы рождается:

Уже с первого шага очевидно, что борьба двух подбираемых форм неравная; во втором поколении рождаемость одной вдвое превышает рождаемость второй, в третьем и четвертом немного менее, чем вдвое, в пятом почти вдвое, а начиная с VI неравенство все возрастает, достигая наконец в X своего maximum'a, именно без малого в пять раз. Вот сравнительная рождаемость, вычисленная сообразно предыдущей таблице:
На 100 особей рано уклонившихся рождается:

Если же возьмем в расчет, что вдобавок существует и абсолютное неравенство размножающих то и другое уклонение особей, то легко поймем быстроту процесса вытеснения одной формы и преобладания другой, хотя обе формы были первоначально равночисленны и равно нравятся и подбираются самками.
Теперь нам надо припомнить те осложняющие влияния некоторых законов изменчивости и наследственности, которыми мы в начале пренебрегли, и рассмотреть, не видоизменят ли они действие закона ограниченной полом наследственности в том упрощенном виде, в котором мы прилагали ее к нашей гипотезе. Переберем все возражения, которые, с этой точки зрения, можно сделать против гипотезы, а следовательно, и против доказательности вычисления. Прежде всего, могут сказать, вы предполагаете в самом начале невозможное, что игра изменчивости вызвала у одних лишь самцов такой значительный % однородных уклонений, как 6 000 из 15 000 родившихся самцов. Конечно, такой % уклонений в действительности невероятен, но уклонение 2-х, 3-х или даже десятка самцов без одновременного уклонения самок весьма возможно, так как самцы вообще значительно изменчивее. Если же это возможно, то, сохраняя остальные условия, результат вычисления останется тот же; только нам для наглядности пришлось бы вычислить не 10, а, быть может, 110 поколений, потому что поздно уклонившиеся продолжали бы размножаться гораздо дольше. С другой стороны, предположение появления уклонений не только у самцов, но и у самок тоже не изменило бы конечного результата вычисления. Поздно уклонившиеся самки вовсе не повлияли бы на него, передавая свои уклонения только самкам и, не будучи подбираемы, вероятно, скоро исчезли бы; рано же уклонившиеся самки только ускорили бы процесс. Следовательно, незаконное преувеличение вероятного числа уклонения самцов от нормального типа оказывается вовсе не таким незаконным, потому что только ускоряет ход процесса, а так как мы наблюдаем только качественную, а не количественную сторону процесса, то такое предположение оказывается вполне целесообразным. Могут еще возразить, что гипотеза равного числа уклонений в раннем и зрелом возрасте тоже не совсем вероятна; с этим нельзя не согласиться отчасти. Изменчивость, действительно, по-видимому, вызывает большее число уклонений в раннем возрасте, чем в зрелые годы. Это можно заключить a priori, сообразив вероятное действие причин изменчивости. Соотносительная изменчивость только следует за другими изменениями и, таким образом, для вопроса значения не имеет. Упражнение и неупражнение частей и органов, вероятно, произведет большее действие в зрелом возрасте. Прямое и определенное действие условий, вызывающее приспособительные процессы, действует равно на взрослых и малолетних, но последние необходимо менее устойчивы, и потому едва ли не наибольшее число уклонений вызовет этот деятель в молодых особях[29]. Реверсия, одна из причин изменчивости, обыкновенно проявляясь при рождении, значительно реже развивается в более позднем возрасте; неопределенная игра изменчивости, вызываемая изменением условий существования или скрещиваньем, действует тремя путями, которым Дарвин придает, по-видимому, одинаковое значение: изменения, претерпеваемые половыми элементами до оплодотворения, изменения зародыша в период утробной жизни, изменения особи уже индивидуализировавшейся – вот эти три разряда уклонений. Первые два, очевидно, произведут уклонения, проявляющиеся уже при рождении, третий же, по причинам, вышеприведенным для определенного действия условий, вероятно, будет деятельнее в молодости, чем в более зрелые годы. Таким образом, едва ли вероятно, чтобы порождалось одинаковое число изменений ранних и поздних; если бы мы предположили, что весь приплод доживет до зрелости, то очень невероятно, чтобы после этого появилось такое же число уклонений, как при рождении и в молодости, а так как мы предполагаем, что естественный подбор безразлично относится к нашему признаку, то тоже нет никакого основания предполагать, чтобы особей с уклонениями ранними погибло бы больше, чем особей, носящих в себе наклонность развить уклонение в зрелом возрасте. Если же это так, то несомненно, что принятое нами условие равного числа ранних и поздних уклонений неверно, но как должна отразиться наша поправка на ходе и характере процесса? Ясно, что она ускорила бы его, не изменяя его характера.
Далее, невероятно, чтобы изменчивость ограничилась только одним поколением, а затем уже предоставила наследственности и половому подбору распределять, стирать или распространять порожденные ею признаки. Раз вызванная к действию, она произведет, конечно, уклонения и во II, III и т. д. поколениях, причем, с одной стороны, она будет в потомках неуклонившихся особей развивать подбираемый признак, с другой – лишать его потомков особей уклонившихся; из этих двух тенденций первая, вероятно, возьмет верх, потому что раз начавшаяся изменчивость имеет наклонность скорее развиваться в ту же сторону, чем в противуположную. Так как, конечно, нет никакого основания предполагать, чтобы в следующих поколениях уклонения распределялись по возрастам иначе, чем в первом (средним счетом они должны распределяться одинаково), то продолженное действие изменчивости будет только мультиплицировать эффекты наследственности и подбора; процесс только ускорится и облегчится, не изменяя ни направления, ни конечного результата. Но наклонность раз начавшейся изменчивости развиваться в том же направлении должна выразиться еще в усилении у потомков уклонившихся особей родительского признака; здесь надо принять во внимание два закона. Во-первых, изменчивость проявляется сильнее в более многочисленных породах, вследствие чего надо предполагать, что рано уклонившиеся индивиды произведут большее число усиленных уклонений, которые, вероятно, установят градацию привлекательности между самими уклонившимися особями. Во-вторых, несколько ослабляет значение этого соображения закон наследственной изменчивости: отцовская изменчивость наследуется сильнее материнской, так что рано уклонившиеся особи произведут потомков с усиленными уклонениями, хотя и больше, нежели поздно уклонившиеся, но непропорционально своему численному превосходству. Во всяком случае, осложнения процесса, порожденные установлением степеней привлекательности, не отразятся на процессе подбора изменением его окончательного результата и даже, по-видимому, будут благоприятны ранним уклонениям и, таким образом, ускорят процесс. Итак, рассмотрев те нарушения законов изменчивости, которые допущены были в нашей гипотезе, мы видим, что введение этих условий только значительно осложнило бы вычисление, не изменив ни конечного результата, ни характера процесса: одни условия отдаляют, другие приближают разрешение, исход борьбы разных пород. Переходим к рассмотрению нарушений законов наследственности.
Самым крупным возражением с этой стороны против доказательной силы вышеприведенных таблиц, несомненно, является закон передачи признаков без ихразвития, так что потомки женского пола поздно уклонившихся особей могут передавать своим сыновьям признаки своих отцов, их дедов. Это несомненно; но весьма важно знать, передаются ли они с такою же силою, как в случае, когда эти признаки не только передаются, но и развиваются матерью. Этого не должно быть, и вот на каких основаниях. Развитие известных признаков у внуков, когда у дочери их не было, принадлежит, конечно, к разряду случаев реверсивных. Это уже сразу должно указать ее меньшую силу, сравнительно с прямым унаследованием. Дарвин говорит[30]: "Обстоятельства, вредящие наследственности, насколько мы их знаем до сих пор, суть … скрещиванье различных разновидностей в каком-либо из предыдущих поколений вместе с реверсией, или атавизмом, т. е. стремлением потомков походить на своих дедов или даже более отдаленных предков, вместо непосредственных родителей". Тут Дарвин рассматривает реверсию только как фактор, колеблющий наследственность. Еще яснее он выражается когда пишет[31]: "В общем итоге можно сказать, что наследственность – правило, изменчивость – исключение. В некоторых случаях известный признак не наследуется потому, что окружающие условия прямо противятся его появлению; в других случаях потому, что эти условия постоянно вызывают новые уклонения… В остальных случаях это отступление может быть приписано возвращению к дедовскому или еще более отдаленному типу". Итак, передача дедовского признака, не развитого самою особью, есть только отступление от общего правила; унаследование же признаков непосредственных родителей и есть это общее правило. Кроме того, реверсия бывает 2 родов: а именно, реверсия, появляющаяся у разновидностей или племен, которые не были скрещены и утратили, вследствие вариации, какой-либо признак, которым обладали прежде и который вновь появляется впоследствии. Второй отдел заключает в себе все те случаи, в которых "известная особь, подразновидность, племя или вид были скрещены в какой-либо прежний период с отличною формою, и признак, заимствованный от этого скрещивания и исчезнувший в течение одного или нескольких поколении, вдруг появляется вновь"[32]. Очевидно, наш случай относится ко второму отделу, про который далее Дарвин говорит[33], что невозможно указать, чрез сколько поколений стремление к реверсии исчезает, но прибавляет, что это рассуждение не должно применять к первому отделу, когда реверсия может проявиться чрез неопределенно долгое время. Ясно, значит, что если и вообще реверсия только фактор частный, исключение из общего правила унаследования родительских признаков, то второй отдел ее как проявляющийся слабее во времени должен рассматриваться как еще более исключительный. То обстоятельство, что этого рода реверсия невозможна после определенного ряда поколений, доказывает, что наследственность от одного родителя имеет наклонность ослабевать с каждым поколением, пока, так сказать, не выветрится совершенно; значит, она должна отчасти ослабеть уже во втором поколении. Из этих соображений следует, что поздние уклонения самцов, не наследующиеся самками, хотя и должны передаваться ими своим сыновьям, но непременно слабее, чем ранние уклонения, которые в самках не только передаются, но и развиваются. Но если это справедливо, т. е. если сила передачи потомков женского пола, происшедших от поздно уклонившихся самцов, слабее силы, с которою самки передают уклонения, унаследованные ими от рано уклонившихся самцов, то окончательный исход процесса будет тот же, как если бы дочери поздно уклонившихся вовсе не передавали бы своим сыновьям признаков своих отцов. В самом деле, мы знаем, что преобладание ранних уклонений в нашем вычислении зависит: 1) от того, что с кем бы ни спарились рано уклонившиеся самки, они произведут потомство рано уклонившееся, даже от самцов поздно уклонившихся, хотя, быть может, в этом случае данный признак будет ярче у детенышей мужеского пола; и 2) от того, что вообще число особей, передающих ранние уклонения, уже со второго поколения больше числа особей, размножающих поздние уклонения. Изменятся ли эти два условия от введения нового соображения? Первое, конечно, не изменится, второе же претерпит изменение только частное, но не будет совершенно парализовано, потому что только часть дочерей поздно изменившихся воспроизведет в своих сыновьях отцовский признак, а с другой стороны, часть этих дочерей спарится с самцами рано уклонившимися и произведет потомков обоего пола рано уклонившихся. Это ведет нас к убеждению, что даже если бы все дочери поздно уклонившихся передавали сыновьям данный признак, то и тогда исход борьбы в пользу ранних уклонений был бы неизбежен; во-первых, потому, что ранние уклонения многочисленнее поздних, во-вторых, даже при гипотезе равного числа уклонений все особи рано уклонившиеся, и мужеского, и женского пола, оставят потомство обоего пола рано уклонившееся, тогда как часть особей, передающих поздние уклонения, произведет потомство обоего пола уклонившееся, хотя, вероятно, не в равной степени. Таким образом, даже при самых невозможно благоприятных условиях, при предположении равночисленности ранних и поздних уклонений, при предположении равенства силы передачи без унаследования и силы непосредственной наследственности, все же оказывается, что раз начавшееся уклонение, подвергаясь действию полового подбора, неизбежно распространяется на оба пола и пренебреженный нами закон передачи признака через дочерей внукам только замедляет процесс, не влияя на его исход.
Мы должны теперь ввести в нашу гипотезу еще два закона наследственности, нами пренебреженные для упрощения задачи. Первый закон: признак, которым обладают оба родителя, передается постояннее и притом стремится усилиться в потомстве. То, что он передается постояннее, важно, ибо доказывает, что унаследование ранних уклонений потомством тех пар, где и отец, и мать уклонившиеся, прочнее унаследования поздних уклонений. Но еще важнее усиленное развитие признака, потому что это устанавливает в среде уклонившихся степени привлекательности; мы видели, что подобная градация вызывается самою изменчивостью и что есть Основание предполагать, что она будет благоприятна ранним уклонениям. Вводимый теперь нами закон наследственности делает это несомненным. Вследствие этого половой подбор, который мы предположили равно благоприятным для ранних и поздних уклонений, вмешается в процесс борьбы двух форм наследственности и склонит победу на сторону ассимиляции обоих полов против их дифференцования. Относительно вопроса, не будет ли спариванье поздно уклонившихся с дочерьми поздно уклонившихся производить такие же последствия? – ответ будет несомненно аналогичный вышесделанному: да, последствия будут такие же, но в меньшей степени как по качеству, так и по количеству.
Второй, опущенный нами закон наследственности – стремление признака развиваться в потомках несколько раньше, чем в родителях и предках. Это, конечно, значительно облегчило бы процесс ассимиляции, если бы не мешала тому трудность перехода (а не вытеснения – это надо различать) одной формы наследственности в другую. Но так как возможность такого перехода отрицать нельзя, то мы можем с некоторой вероятностью заключить, что, сочетаясь с другими вышепоименованными факторами, этот закон может содействовать ускорению процесса, который произошел бы и без него. Итак, все рассуждения ведут нас к тому, что наше вычисление соответствует вообще характеру процесса, который совершился бы при взаимодействии всех условий, нами опущенных и измененных. Восстановление одних из них замедляет, введение других ускоряет ход процесса, но ни один из опущенных деятелей не изменяет исхода процесса, так что мы приходим к тому конечному выводу, что половой подбор, предоставленный самому себе, вообще стремится изменить всю породу без различия пола и даже, пожалуй, ведет скорее к ассимиляции, чем к дифференцованию полов.
Дарвин держится, по-видимому, другого воззрения[34]: "Возьмем пару животных, не особенно плодовитых, не особенно бесплодных, и предположим, что они живут средним числом 5 лет, по достижении половой зрелости, производя ежегодно по 5 детенышей. Они произведут, таким образом, потомство в 25 штук; и я полагаю, что немного удалюсь от истины, если сделаю предположение, что из 25 неделимых погибнет 18 или 20 штук, не достигнув зрелости, в период юности и неопытности; остающихся же 7 или 5 неделимых будет достаточно для поддержания колена зрелых особей. При таком предположении легко видеть, что изменения, случившиеся в юности, напр. в отношении красоты и не приносящие молодому животному никакой выгоды, будут иметь шансы быть утраченными впоследствии. Наоборот, подобные же изменения, появившиеся во время или около зрелости на сравнительно меньшем числе неделимых, доживших до этого возраста, которые непосредственно приносят пользу самцам, делая их более привлекательными для самок, легко сохраняются". Ошибочность этого рассуждения, после всего вышеизложенного, очевидна. Во-первых, если не принять во внимание естественного подбора (которого Дарвин в этом месте не берет в расчет), то нет оснований предполагать, что расположенных к изменчивости в зрелости выживет больше, чем изменившихся в юности, а между тем Дарвин это подразумевает, и, во-вторых, что касается полезности украшений зрелым самцам и бесполезности молодым, то едва ли по этому поводу можно говорить о влиянии неупражнения, вследствие чего они, дескать, могут их и потерять, потому что ведь и взрослые, хотя и пленяют ими самок, однако трудно это пленение назвать упражнением. Относительно вооружения с этим еще можно бы было согласиться, если бы молодежь не упражняла их в играх. Во всяком случае, впрочем, эта небольшая разница, если и существует, покрывается с лихвою большим количеством ранних уклонений, и мы, быть может, должны прибавить одну причину к списку деятелей, замедляющих неизбежный исход полового подбора, когда он предоставлен самому себе. Сам Дарвин отказался, по-видимому, от вышеприведенного аргумента[35], но все же в продолжении всего сочинения поддерживает мнение, что именно половой подбор производит вторичные половые признаки, и постоянно оспаривает мнение Уоллеса о естественном подборе как причине дифференцования полов. Это нас ведет к рассмотрению вопроса о влиянии естественного подбора на половой подбор, вопроса, выше нами отложенного для простоты задачи. Дарвин высказывает следующий взгляд на значение естественного подбора как начала, осложняющего эффекты полового подбора: "Многие из признаков, свойственных взрослому самцу, были бы положительно вредны молодым, так как яркая окраска делала бы их более видными для глаз неприятеля, большие рога требовали бы большего расхода сил"[36]. Вследствие этого естественный подбор имеет общую тенденцию уничтожать особей, приобретающих в раннем возрасте какие-либо особые украшения или вооружения, потому что если вооружения и полезны взрослой особи не только в соперничестве из-за самок, но и в борьбе за существование, то для молодых особей они не могут принести пользы в борьбе за существование по их слабости, а между тем закон экономии роста дает преимущество особям, не имеющим их. Таким образом, часто естественный подбор действует вовсе не безразлично на признаки, подверженные половому подбору; благодаря его влиянию, поздние уклонения получают громадное преимущество, особенно у пород, истребляемых многочисленным неприятелем. Но этим истреблением в молодости рано уклонившихся особей не ограничивается естественный подбор; он часто и в зрелом возрасте преследует уклонения, подбираемые самками. Правда, относительно самцов здесь равно гибнут оба разряда особей, с поздними и ранними уклонениями, и эта гибель равно вознаграждается половым подбором, но относительно самок этого сказать нельзя: 1) потому, что передающие поздние уклонения самки остаются сами без уклонений и не преследуются естественным подбором и 2) потому, что гибель самок не вознаграждается их подбором. Большая слабость самки, критические периоды вынашиванья и выкармливанья детенышей составляют новый ряд фактов, способствующих устранению всяких невыгодных для жизни уклонений у самок. Этим путем совершенно видоизменяется характер и исход процесса полового подбора, и получается в результате дифференцование полов, вторичные половые признаки. Ни естественный подбор сам по себе, ни половой подбор независимо от естественного не произвели бы этого дифференцования, но их сложное действие часто приводит к этому неизбежным путем, именно: во всех случаях, когда признак, распространяемый половым подбором, устраняется естественным подбором. Тот же результат должен произойти, если бы признак, распространяемый половым подбором, искоренялся прямым действием условий жизни.
Влияние прямого действия условий жизни на ход и исход полового подбора тем важно, что оно может произвести дифференцование полов не только противодействуя, но и содействуя половому подбору, именно: когда чрез упражнение, вынужденное новыми условиями жизни, какая-нибудь часть организма получает большее развитие, и эта гиперестезия понравится самкам. Может случиться, что особые условия жизни самцов произведут и другие изменения самцов только в зрелом возрасте, а половой подбор распространит их.
Итак, резюмируя наше предварительное рассуждение о половом подборе вообще, мы можем сказать: половой подбор зависит от взаимодействия законов изменчивости, наследственности и полового инстинкта, управляемого эстетическим вкусом, при условии неравночисленности полов, абсолютной или только условной, как при полигамии или неединовременном созревании самок. Последнее условие есть условие только отрицательное, и та или другая форма его не влияет на процесс, но присутствие которой-либо из них необходимо; иначе половой подбор не может начать своего действия. Вообще половой подбор имеет наклонность изменять всю породу без различия пола; предоставленный самому себе, он не ведет к дифференцованию полов. Этот результат получается, когда он сталкивается в своем развитии с естественным подбором или прямым влиянием условий; в том и другом случае чрез устранение ранних уклонений, а также женских уклонений получает преобладание наследственность, ограниченная полом. К тому же результату может привести и согласное действие условий и полового подбора, именно: когда условия влияют только на один пол и в зрелом возрасте. Впрочем, в этом последнем случае, дифференцованние только ускоряется половым подбором, а производится прямым, определенным действием условий.