Углеводы
Углеводы широко представлены в растениях и животных, где они выполняют как структурные, так и метаболические функции. В растениях в процессе фотосинтеза из углекислого газа и воды синтезируется глюкоза, которая далее запасается в виде крахмала или превращается в целлюлозу – структурную основу растений. Животные способны синтезировать ряд углеводов из жиров и белков, но большая часть углеводов поступает с пищей растительного происхождения.
Классификация углеводов
Углеводы – это альдегидные или кетонные производные полиатомных (содержащих более одной ОН-группы) спиртов или соединения, при гидролизе которых образуются эти производные.
Их можно классифицировать следующим образом:
Моносахариды – углеводы, которые не могут быть гидролизованы до более простых форм. Их можно подразделить на триозы, тетрозы, пентозы, гексозы, гептозы и октозы в зависимости от числа содержащихся в их молекуле атомов углерода; их можно разделить также на альдозы и кетозы в зависимости от присутствия альдегидной или кетонной группы.
Дисахариды при гидролизе дают две молекулы моносахарида (одинаковых или различных).
Олигосахариды при гидролизе дают 3—6 моносахаридов или, что чаще, полисахариды, в состав которых входит несколько десятков моносахаридных остатков.
Полисахариды дают при гидролизе более 6 молекул моносахаридов. Они могут быть линейными или разветвленными. Если полисахарид состоит из одинаковых мономеров, то их их называют гомополисахаридами, если из разных, то гетерополисахаридами. Также следует отметить, что большая часть полисахаридов содержит более тысячи мономерных остатков. Примерами служат крахмал и декстрины.
Моносахариды
Моносахариды – углеводы, которые не могут быть гидролизованы до более простых форм. Их можно подразделить на триозы, тетрозы, пентозы, гексозы, гептозы и октозы в зависимости от числа содержащихся в их молекуле атомов углерода; их можно разделить также на альдозы и кетозы в зависимости от присутствия альдегидной или кетонной группы. Отсчет атомов углерода начинают либо от входящего в состав альдегидной группы (первый атом), либо от ближайшего к кетонной группе.
Изомерия моносахаридов
Соединения, имеющие одну и ту же структурную формулу, но различающиеся по пространственной конфигурации, называются изомерами. Образование таких изомеров оказывается возможным при вхождении в состав молекулы асимметрических атомов углерода (к которым присоединены четыре различных атома или группы). Число возможных изомеров данного соединения зависит от числа асимметрических атомов углерода (n) и равно 2n. Глюкоза с четырьмя асимметрическими атомами углерода имеет, следовательно, 16 изомеров. Ниже указаны наиболее важные типы изомеров глюкозы.
Стереоизомерия или D и L изоформы:
В многих органических молекулах есть атом углерода с которым связаны четыре различные группировки, и такую молекулу можно представить как молекулу метана, в виде тетраэдра, в верхушках которого располагаются группировки. Такой атом называется хиральным, в случае моносахаридов принадлежность к D или L форме определяется расположением гидроксильной (ОН) группировки, относительно хирального атома углерода. Если ОН-группа слева, то это L-форма, если справа, то D-форма. Но полностью этому правилу соответствует только глицероальдегид, имеющий только один хиральный атом (Рисунок 1 А). Во всех других углеводах хиральных атомов несколько, и каждый из них может быть признан для определения принадлежности к одной из форм стереоизомеров. Было принято решение признать атомом, определяющим стереоизомерию, последний хиральный атом молекулы (предпоследний атом углерода в молекуле). Но изменение положения гидроксильной группы только у данного атома углерода не приведет к образованию полностью зеркального отображения, что является основным условием для существования двух форм стереоизомеров. Поэтому появилось понятие ряда углеводов, то есть все углеводы могут быть выведены или синтезированы из минимального углевода (триозы), и, следовательно, эта триоза является предшественником или родительским соединением, так как единственная триоза, обладающая оптическими свойствами – глицероальдегид, то именно эта молекула является родительским соединением для всех остальных. Принадлежность к одному их стреоизомеров родительского соединения, определяет принадлежность к D или L-форме всех остальных моносахаридов, синтезируемых из данного родительского соединения (Рисунок 2). В ходе синтеза углеводов с числом атомов больше чем три (глицероальдегид), присоединение каждого следующего атома углерода

Рисунок 1. Структурные формулы стереоизомеров углеводов, * – отмечены асимметрические (хиральные) атомы. А – глицероальдегид * – отмечен асимметрический атом, определяющий принадлежность к D или L форме, Б – стереоизомеры глюкозы, крупной * отмечены асимметрические атомы углерода, определяющие принадлежность к стереоизомерам
происходит по альдегидной группе, и, следовательно, хиральный атом родительского соединения оказывается все дальше от первого атома углерода, то есть остается последним асимметричным или просто предпоследним атомом углерода (рисунок 2). Присутствие асимметрических атомов углерода является причиной оптической активности соединения. Если пучок плоскополяризованного света проходит через раствор оптического изомера, плоскость поляризации света поворачивается либо вправо (правовращающий изомер, +), либо влево (левовращающий изомер, – ). Соединение обозначают D ( – ), D (+), L ( – ) или L (+); это обозначение показывает наличие структурного родства с D- или L- глицеральдегидом, но не обязательно тот же знак оптического вращения. Например, природной формой фруктозы является D ( – ) – изомер. Если D- и L-изомеры присутствуют в равных количествах, их смесь не проявляет оптической активности – активности изомеров компенсируют одна другую. Такие смеси называют рацемическими (или DL-смесями). Соединения, получаемые синтетическим путем, оказываются рацемическими, поскольку в этом случае вероятности образования каждого из изомеров одинаковы.

Рисунок 2. Структура D-ряда углеводов
2. Пиранозные и фуранозные кольцевые структуры.
Альдегидные или кетонные группировки легко реагируют со спиртовыми, образуя полуацетальные или полукетальные группировки, чаще всего это происходит внутри молекулы, и, следовательно, происходит циклизация моносахарида, в результате образуется модифицированный гетероцикл, содержащий атом кислорода. Наиболее устойчивы пяти и шестичленные циклы. Пятичленные циклы углеводов сходны с молекулой фурана, поэтому это фуранозная форма, шестичленные с молекулой пирана – пиранозная форма (Рисунок 3). Все гидроксильные группы, расположенные справа оказываются под циклом, а те что слева – над циклом. Кольцевую структуру могут принимать и кетозы (например, D-фруктофураноза или D-фруктопираноза). В растворе глюкозы более 99% молекул находится в пиранозной форме и менее 1% – в фуранозной форме.

Рисунок 3. Пиранозные и фуранозные формы углеводов
3. α- и β-аномеры. При циклизации образуется гидроксильная группа при полуацетальной или полукетальной группировке, полуацетальный или полукетальный гидроксил, эта группа может располагаться под циклом, в результате образуется α-аномер, а может располагаться над циклом, тогда образуется β-аномер. Циклическая структура сохраняется и в растворе, но при этом происходит образование изомеров относительно положения альдегидного и кетонного атома углерода, что приводит к образованию смеси α – глюкопиранозы (36%) и β – глюкопиранозы (63%); оставшийся 1% представлен в основном α – и β – аномерами глюкофуранозы. Описанное выше установление равновесия сопровождается так называемой мутаротацией: полуацетальное кольцо раскрывается и вновь замыкается, при этом может изменяться положение групп – Н и – ОН при углероде 1. Предполагают, что в ходе этого процесса образуется промежуточная гидратированная линейная (ациклическая) молекула, хотя по данным полярографии на долю ациклической формы глюкозы приходится всего 0,0025%. В растворе глюкоза является правовращающей; этим объясняется еще одно ее название – декстроза (декстро – правый), часто употребляемое в клинической практике (Рисунок 4).

Рисунок 4. Структурные формулы аномеров глюкозы. α- и β-аномеры
4. Эпимеры. Изомеры, различающиеся по конфигурации положением групп – Н и – ОН при асимметричных атомах углерода, которые не связаны с принадлежностью к стереоизомерам, называются эпимерами. Биологически наиболее важными эпимерами глюкозы являются: манноза и галактоза, образующиеся путем эпимеризации при атомах углерода 2 и 4 соответственно (Рисунок 5).

Рисунок 5. Эпимеризация глюкозы
5. Альдо-кето-изомеризация. Фруктоза имеет ту же химическую формулу, что и глюкоза, но отличается по структурной формуле, поскольку фруктоза содержит потенциальную кетонную группу в положении 2, а глюкоза – потенциальную альдегидную группу в положении 1.
Конформация моносахаридов
Шестичленные кольца из одинарных связей, в циклогексане и в сахарах, как правило, принимают конформацию «кресла», как это видно на примере глюкозы (Рисунок 6). Помимо этого, шестичленные кольца могут принимать менее устойчивую конформацию «лодки». Свободно переходят одна в другую через промежуточные скошенные конформации.
Пятичленные кольца образуют конформацию «конверта».

Рисунок 6. Конформация шестичленных колец
Физические свойства моносахаридов
Моносахариды гидрофильны, а, следовательно, легко растворимы в воде. Растворимость моносахаридов примерно одинакова, поэтому разделение моносахаридов достаточно сложно. Моносахариды не поглощают свет ни в видимой, ни в ультрафиолетовой части спектра. Из-за этого нельзя определять концентрацию моносахаридов спектрофотометрическими методами. Так как моносахариды находятся в природе либо в растворе, либо в кристаллах, в физиологических диапазонах температуры нет вопроса о температурах плавления и кипения.
Химические свойства моносахаридов
По своей структуре моносахариды содержат два типа функциональных групп: спиртовые и альдегидные (кетонные), соответственно моносахариды вступают в реакции по этим группам. Моносахариды вступают во многие химические реакции, весьма важные для практического использования и исследования углеводов. Реакции, обсуждаемые в данном разделе, демонстрируют либо наиболее важные свойства углеводов, либо типы реакций, часто встречающиеся в их метаболизме.
Образование эфиров. Это реакция спиртовых групп образуются либо эфиры неорганических кислот, либо органические эфиры (простые со спиртами или галогенидами или сложные с ангидридами кислот). Наиболее активной является полуацетальная спиртовая группа, соответственно она чаще вступает в реакции с образованием эфиров. Эфиры фосфорной и уксусной кислот занимают уникальное место в биохимии и встречаются во многих углеводах и их производных.
Алкилирование (Рисунок 7)
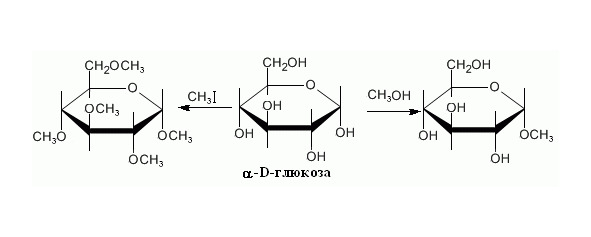
Рисунок 7. Алкилирование
При взаимодействии со спиртами в реакцию вступает только полуацетальная гидроксильная группа, так как спирты слабо активны, поэтому образуется эфир с полуацетальной гидроксильной группой. Галогениды более реакционно способны, поэтому образуют эфиры со всеми свободными группами моносахарида.
Ацилирование (Рисунок 8)

Рисунок 8. Ацилирование
В реакцию образования сложных эфиров вступают спиртовые и карбоксильные группы, но карбоновые кислоты мало реакционно способны поэтому не вступают в реакцию моносахаридами. В реакции ацилирования используют ангидриды карбоновых кислот, их высокая реакционная способность позволяет модифицировать все свободные гидроксильные группы.
Образование эфиров неорганических кислот (Рисунок 9)

Рисунок 9. Образование эфиров неорганических кислот
Неорганические кислоты бывают сильными и слабыми, это объясняет различия их взаимодействия с моносахаридами. Сильные кислоты, например серная, вступают в реакцию с полуацетальной гидроксильной группой, образуя эфир, слабые кислоты вступают в реакцию только после реакции ацилирования, но модифицируют только эфир образованный полуацетальной группой.
Дегидратация. В сильных минеральных кислотах высокой концентрации происходит дегидратация пентоз и гексоз (Рисунок 10). Образовавшиеся фурфурол и оксиметилфурфурол легко полимеризуются; при этом получается коричневая смола. Они также конденсируются с различными фенолами, образуя характерно окрашенные продукты; многие цветные реакции на углеводы основаны на этом свойстве.

Рисунок 10. Дегидратация
Перегруппировка в щелочной среде. В холодном разбавленном щелочном растворе глюкоза образует маннозу и фруктозу. Механизм реакции, возможно, включает енолизацию, которая сопровождается диссоциацией водорода от атома углерода, примыкающего к карбонильной группе. Так образуются эпимеры – глюкоза и манноза.
Окисление моносахаридов. Окисление моносахаридов, возможно, многими способами.
Мягкое окисление до альдоновых кислот. В горячих щелочных растворах моносахариды реагируют с некоторыми окисляющими агентами, такими, как Cu2+, Ag+ и Fe (CN) -, давая смесь продуктов окисления углевода и изменяя окраску окисляющего агента (Рисунок 11). В данных условиях окисляется альдегидная или кетонная группа. Эти окислительно-восстановительные реакции используются как тесты на восстанавливающие сахара, т. е. на углеводы с незамещенным аномерным альдегидным атомом углерода. D-Глюконовая кислота образуется в виде соли при взаимодействии D-глюкозы с гипоиодитами в щелочной среде. Эта реакция специфична для альдоз и используется для того, чтобы отличить их от кетоз.

Рисунок 11. Мягкое окисление моносахаридов до альдоновых кислот
Более жесткое окисление до альдуроновых кислот, реакция происходит при участии «средних» окислителей, например небольшие концентрации азотной кислоты (Рисунок 12). Данная реакция происходит и в природе, но при других условиях. В результате окисляется гидроксильная группа при последнем атоме углерода.

Рисунок 12. Окисление до альдуроновых кислот
Жесткое окисление до альдаровых кислот, реакция происходит в жестких условиях, например при больших концентрациях сильных кислот, например, азотной. В результате происходит окисление обеих групп – альдегидной и последней спиртовой, в результате образуется дикарбоновая кислота, или альдаровая (Рисунок 13).

Рисунок 13. Окисление до альдаровых кислот
Восстановление моносахаридов. Восстановлению могут подвергаться как альдегидная, так и спиртовые группы
Восстановление альдегидной группы. D-Глюкоза и L-сорбоза восстанавливаются газообразным водородом в присутствии подходящего металлического катализатора, образуя сорбит (Рисунок 14).
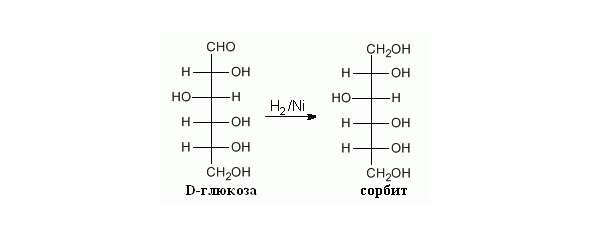
Рисунок 14. Восстановление альдегидной группы
Восстановление спиртовой группы. В клетке происходит переход рибозы в дезоксирибозу (Рисунок 15). Эта реакция происходит в природе, но при несколько иных условиях. Это один из основных способов синтеза дезоксисахаров.

Рисунок 15. Восстановление спиртовой групы
Образование аминосахаров. В природе также происходит синтез аминосахаров, но механизм и атакуемые группы отличаются, в искуственной системе в реакцию вступает полуацетальная группировка как наиболее реакционно способная (Рисунок 16).
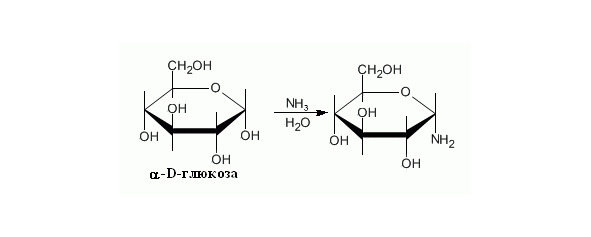
Рисунок 16. Образование аминосахаров
Укорочение цепи. Этот процесс связан с окислением в средних условиях. Карбоксильная группа в последнем положении нестабильна и легко отщепляется от молекулы, в результате углевод становится на один атом короче (Рисунок 17).

Рисунок 17. Укорочение цепи
Удлинение цепи. Это результат реакции с цианидом, затем происходит реакция с водой, в результате образуется кислота. Восстановление карбоксильной группы приводит к образованию моносахарида, но на один атом углерода длиннее (Рисунок 18).
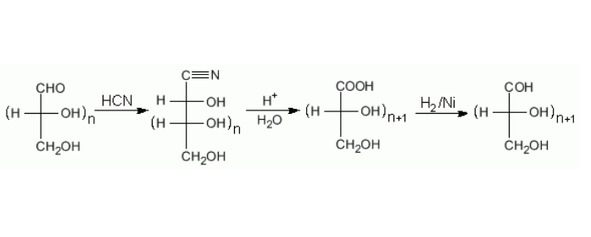
Рисунок 18. Удлинение цепи
Гидролиз поли- и олигосахаридов. Реакция гидролиза это реакция расщепления полимера с участием молекулы воды. В случае гидролиза поли- и олигосахаридов атаке подвергается гликозидная связь. В результате реакции отщепляются мономерные остатки (моносахариды). Реакции гидролиза поли- и олигосахаридов с последующим анализом продуктов гидролиза используются для качественного и количественного определения состава полисахаридов.
Определение моносахаридов
Некоторые качественные реакции, которые используются для идентификации моносахаридов.
Альдозы, кетозы
Реактив: α-нафтол (реакция Молиша), триптофан аминогуанин.
Более чувствительна для кетоз.
Кетогексозы
Реактив: резорцин (реакция Селиванова).
Кетогексозы, Кетопентозы, метилпентозы, диоксиацетон
Реактив: цистеин/карбазол.
Все углеводы, включая уроновые кислоты и дезоксипентозы
Реактив: карбазол.
Характерное окрашивание со всеми углеводами.
Многие углеводы, включая полисахариды, чаще используется для гексоз.
Реактив: цистеин/H2SO4, антрон.
Неодинаковое окрашивание с различными углеводами.
Пентозы, гептулозы, уроновые кислоты
Реактив: орцин.
Окрашивание, обусловленное присутствием других углеводов, может быть ликвидировано независимыми методами, уроновые кислоты декарбоксилируются до пентоз и вступают в реакцию.
Уроновые кислоты
Реактив: нафтилрезорцин.
гексозамины
Реактив: Ацетилацетон-n-диметил-аминобензальдегид.
гексозамины
Реактив: нитрит/индол.
Аминосахара не дают окрашивания без предварительного дезаминирования нитритом.
Моно и дидезоксипентозы
Реактив: дифениламин.
дезоксипентозы
Реактив: триптофан/HClO4, индол/HCl, лейкофуксин (реакция Фольгена).
Сиаловые кислоты
Реактив: тиобарбитуровая кислота.
Функции моносахаридов
Функции моносахаридов очень разнообразны и зависят от того сколько атомов углерода содержит моносахарид.
Триозы – промежуточные продукты обмена углеводов и липидов.
Тетрозы – промежуточные продукты обмена углеводов могут входить в состав полисахаридов.
Пентозы – промежуточные продукты обмена углеводов могут входить в состав полисахаридов, и нуклеотидов.
Гексозы – глюкоза и фруктоза основные сахара энергетического обмена углеводов, входят в состав полисахаридов.
Гептозы – промежуточные продукты обмена углеводов.
Физиологически важные моносахариды
D-рибоза
В какие молекулы или вещества входит: нуклеотиды, коферменты, РНК.
Биологическое значение: компонент нуклеиновых кислот коферментов (NAD, NADP, FAD), нуклеотидов, промежуточное соединение пентозофосфатного пути.
D-рибулоза
В какие молекулы или вещества входит: образуется в ходе метаболизма.
Биологическое значение: промежуточное соединение пентозофосфатного пути.
D-Арабиноза
В какие молекулы или вещества входит: гуммиарабик, сливовая и вишневая мякоть.
Биологическое значение: компонент гликопротеинов.
D-Ксилоза
В какие молекулы или вещества входит: древесная смола, протеогликаны, гликозаминогликаны.
Биологическое значение: компонент гликопротеинов.
D-Ликсоза
В какие молекулы или вещества входит: ликсофлавин.
Биологическое значение: компонент ликсофлавина, выделяемого сердечной мышцей.
L-Ксилулоза
В какие молекулы или вещества входит: промежуточный продукт метаболизма уроновых кислот.
Биологическое значение: промежуточный продукт метаболизма уроновых кислот.
D-глюкоза
В какие молекулы или вещества входит: фруктовые соки, крахмал, сахароза, лактоза, мальтоза
Биологическое значение: сахар организма, участвует в энергетическом обмене, является предшественником других соединений.
D-фруктоза
В какие молекулы или вещества входит: мед, сахароза, лактоза, инулин.
Биологическое значение: превращается в глюкозу, и может использоваться в тех же метаболических путях.
D-галактоза
В какие молекулы или вещества входит: лактоза, гликопротеины, гликолипиды.
Биологическое значение: превращается в глюкозу, и может использоваться в тех же метаболических путях.
Производные моносахаридов
Производные моносахаридов: эфиры моносахаридов, альдуроновые кислоты, аминосахара, дезоксисахара, гликозиды.
Все производные моносахаридов входят в состав полисахаридов. По мимо этой основной функции производные моносахаридов могут выполнять свои специфические функции.
Альдуроновые кислоты участвуют в образовании витамина С и в процессах детоксикации.
Дезоксисахара входят в состав нуклеотидов (мономеры ДНК).
Аминосахара входят в состав антибиотиков.
Гликозиды
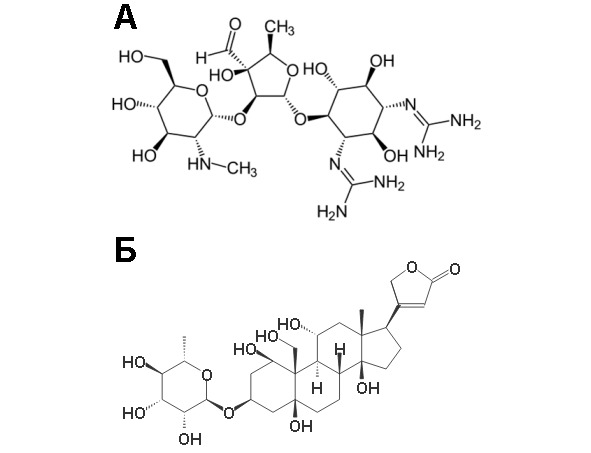
Рисунок 19. Структура некоторых гликозидов. А-стрептомицин, Б-уабаин
Гликозиды – это соединения, образующиеся путем конденсации моносахарида (или моносахаридного остатка в составе более сложного сахара) с гидроксильной группой другого соединения, которым может быть другой моносахарид или вещество неуглеводной природы (тогда его называют агликоном). Гликозидная связь образуется в результате реакции полуацетальной (полукетальной) группы моносахарида и спиртовой группой другого соединения, такая связь называется О-гликозидная. Также полуацетальная (полукетальная) гидроксильная группа может вступать в реакцию с аминогруппой другого соединения тогда образуется N-гликозидная связь. Если полуацетальная группа принадлежит глюкозе, образующееся соединение называют глюкозидом, если галактозе – галактозидом и т. д. Гликозиды найдены в составе многих лекарств и пряностей, они являются также компонентами животных тканей. Агликонами могут быть метанол, глицерол, какой-либо стерол или фенол. Гликозиды, имеющие важное медицинское значение, например, влияющие на работу сердца (сердечные гликозиды), содержат в качестве агликонового компонента стероиды; так, из наперстянки и строфанта выделен гликозид уабаин – ингибитор Na/K-ATP-aзы клеточных мембран. К числу гликозидов относится ряд антибиотиков, в частности стрептомицин (Рисунок 19). N-гликозидами являются нуклеотиды и нуклеозиды. Но простейшими гликозидами являются дисахариды. В случае дисахаридов агликоном является молекула другого моносахарида.
Дисахариды
Дисахариды состоят из двух ковалентно связанных друг с другом моносахаридов. У дисахаридов химическая связь между моносахаридными единицами называется гликозидной связью; она образуется в результате взаимодействия гидроксильной группы одного из сахаров с аномерным атомом углерода второго сахара. Гликозидные связи легко гидролизуются кислотами, но устойчивы к действию оснований. Поэтому дисахариды можно гидролизовать и получить в свободном виде их моносахаридные компоненты путем кипячения в разбавленной кислоте.
По составу дисахариды подразделяют на гомодисахариды (состоят из одинаковых мономеров) и гетеродисахариды (в состав входят разные моносахариды).
По наличию свободной полуацетальной группы дисахариды можно разделить на две группы: восстанавливающие (редуцирующие) и невосстанавливающие (нередуцирующие). Свободная полуацетальная группа способна разделяться с образованием спиртовой и альдегидной или кетонной группы, то есть происходит разрыв цикла. Образовавшаяся альдегидная (кетонная) группировка может вступать реакцию с ионами металлов (Cu2+, Ag+) и восстанавливать их – редуцирующие дисахариды. Если все свободные полуацетальные группировки образуют гликозидные связи, то разрыв цикла невозможен, также как и восстановление альдегидной (кетонной) группы, следовательно не может вступать в реакцию восстановления с ионами металлов – нередуцирующие дисахариды.
Все дисахариды являются гликозидами. Называя дисахарид необходимо учитывать факт, что это гликозиды. Кроме того, при номенклатуре дисахаридов необходимо учитывать также химическую связь между моносахаридами. То есть указываются номера атомов углерода, спиртовые группы при которых участвуют в образовании гликозидной связи, а также аномер моносахарида, чей полуацетальный гидроксил образует гликозидную связь.
Практически все дисахариды выполняют транспортную функцию, так как они легко растворимы, и при этом менее функционально активны чем моносахариды.
Часть дисахаридов, например мальтоза являются промежуточными продуктами гидролиза полисахаридов.
Дисахариды можно подразделить на дисахариды растений и дисахариды животных.
Дисахариды животных
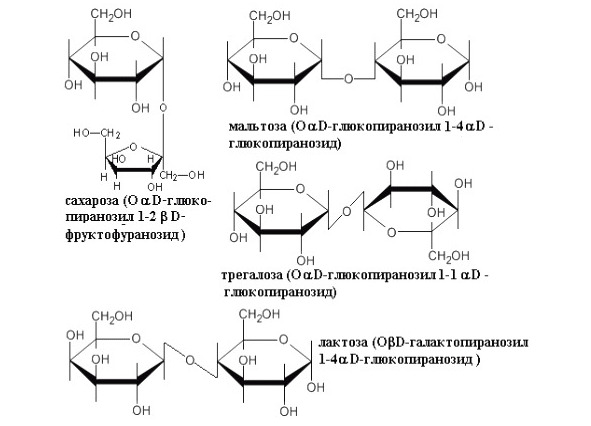
Рисунок 20. Структурные формулы основных дисахаридов
Мальтоза гомодисахарид, содержащий два остатка α D-глюкозы, соединенных α (1—4) гликозидной связью, то есть связь между гидроксилом при 1 углероде и гидроксилом при 4 углероде, причем полуацетальный гидроксил в α форме (Рисунок 20). Оба остатка глюкозы в молекуле мальтозы находятся в пиранозной форме. Мальтоза относится к восстанавливающим сахарам, поскольку она содержит одну потенциально свободную альдегидную группу, которая может быть окислена. Второй остаток глюкозы в молекуле мальтозы может существовать как в α-, так и в β-форме; мальтоза образуется при действии на крахмал содержащегося в слюне фермента – амилазы. Под действием секретируемого слизистой кишечника фермента мальтазы, специфически гидролизующего α (1—4) – связь, мальтоза подвергается гидролизу с образованием двух молекул D-глюкозы.
Лактоза гетородисахарид, при гидролизе которой образуется D-галактоза и D-глюкоза, моносахариды соединены β (1—4) гликозидной связью, присутствует только в молоке (Рисунок 20). Наличие в молекуле лактозы потенциально свободной карбонильной группы (в остатке глюкозы) делает ее восстанавливающим дисахаридом. Лактоза является молочным сахаром, это единственный дисахарид, синтезируемый млекопитающими.
В процессе переваривания пищи лактоза подвергается ферментативному гидролизу в результате воздействия лактазы, секретируемой мукозными клетками кишечника. У грудных младенцев активность этого фермента очень высока, однако в кишечнике взрослых людей лактазная активность наблюдается лишь у жителей севера Европы и некоторых африканских племен. У большинства взрослых людей, в том числе у жителей Востока, арабов, евреев, многих африканцев, индийцев и жителей Средиземноморья, лактазная активность в кишечнике очень низка, что часто приводит к непереносимости (интолерантности) лактозы. Описанная особенность обусловлена генетически. Причина непереносимости лактозы связана с тем, что этот дисахарид может всасываться в кишечнике только после гидролиза на моносахаридные компоненты: при низкой лактазной активности неусвоенная лактоза накапливается в кишечнике; в результате после потребления молока у человека с непереносимостью лактозы возникает тяжелый понос и боли в животе.
Трегалоза состоит из двух молекул α D-глюкозы, соединенных 1—1 α гликозидной связью. Трегалоза входит в состав гемолимфы насекомых, также выделяется из некоторых грибов. Является нередуцирующим дисахаридом (Рисунок 20). Трегалоза является транспортной формой моносахаридов в кровеносной системе насекомых.
Дисахариды растений
Сахароза – гетеродисахарид, состоящий из глюкозы и фруктозы, соединенных β (1—2) гликозидной связью (Рисунок 20). Сахароза является нередуцирующим сахаром. Сахарозу синтезируют многие растения, у высших же животных она отсутствует. В отличие от мальтозы и лактозы у сахарозы нет свободного аномерного атома углерода, поскольку оба аномерных атома моносахаридных остатков связаны друг с другом; поэтому сахароза не является восстанавливающим сахаром. Сахароза основной промежуточный продукт фотосинтеза. У многих растений именно в форме сахарозы транспортируются по сосудистой системе сахара из листьев к другим частям растения. Преимущество сахарозы перед глюкозой как транспортной формы сахаров заключается, вероятно, в том, что ее аномерные атомы углерода связаны друг с другом: это предохраняет сахарозу от атаки окислительных или гидролитических ферментов в процессе ее переноса из одной части растений в другую. Животные не могут усваивать сахарозу как таковую, однако она становится доступной для усвоения после воздействия фермента сахаразы (другое его название – инвертаза), локализованного в клетках, выстилающих тонкий кишечник. Этот фермент катализирует расщепление сахарозы на D-глюкозу и D-фруктозу, которые легко проникают в кровоток.
Полисахариды
В природе большинство углеводов представлено в виде полисахаридов с высокой молекулярной массой. Биологическое значение ряда полисахаридов состоит в том, что одни обеспечивают накопление моносахаридов для энергетического обмена в нерастворимой, а значит осмотически неактивной форме, другие же служат структурными элементами клеточных стенок и соединительной ткани. При полном гидролизе под действием кислоты или специфических ферментов полисахариды расщепляются с образованием моносахаридов или их производных.
Полисахариды, называемые также гликанами, отличаются друг от друга как природой составляющих их моносахаридных остатков, так и длиной и степенью разветвленности цепей. Их можно разделить на два типа: гомополисахариды, состоящие из остатков одного и того же моносахарида, и гетерополисахариды, содержащие остатки двух или большего числа моносахаридов. Пример гомополисахарида резервный углевод крахмал, состоящий из остатков только D-глюкозы. Примером гетерополисахарида может служить содержащаяся в соединительной ткани гиалуроновая кислота, которая состоит из чередующихся остатков двух разных моносахаридов.
В отличие от белков полисахариды нельзя характеризовать строго определенной молекулярной массой: как правило, они представлены смесями высокомолекулярных соединений; в зависимости от метаболических потребностей клеток моносахаридные остатки могут ферментативно присоединяться к полисахаридам или же отщепляться от них. Также, как и дисахариды, полисахариды делятся на редуцирующие и нередуцирующие. По наличию свободной альдегидной группы, которая, окисляясь, восстанавливает ионы некоторых металлов.
По функции полисахариды делят на структурные и запасающие.
Запасающие полисахариды обеспечивают накопление моносахаридов, участвующих в энергетическом обмене в виде компактных нерастворимых структур (включений). Нерастворимость обеспечивает отсутствие влияния на осмотическое давление в клетке.
Структурные полисахариды служат внеклеточными опорными элементами в стенках клеток одноклеточных микроорганизмов, грибов и высших растений, а также входят в состав соединительной ткани позвоночных и экзоскелета членистоногих. Структурные полисахариды защищают клетки, ткани и органы, придают им форму и поддерживают ее. У различных организмов запасающие и структурные полисахариды различаются.
Запасающие полисахариды животных и грибов

Рисунок 21. Структура гликогена
Гликоген – полисахарид, в виде которого углеводы запасаются в организме животного. Его часто называют животным крахмалом. В наибольшем количестве гликоген содержится в печени, где на его долю приходится до 7% общего веса органа; гликоген имеется также в скелетных мышцах. В клетках печени гликоген присутствует в виде крупных гранул, состоящих в свою очередь из меньших гранул; последние образованы единичными сильно разветвленными молекулами гликогена со средней молекулярной массой в несколько миллионов. С этими же гранулами прочно связаны ферменты, ответственные за синтез и распад гликогена. Гликоген откладывается в виде гранул в цитоплазме клетки.
У грибов гликоген запасается в клетках гифов.
Гликоген – редуцирующий гомополисахарид, образованный остатками α-D-глюкопиранозы. Гликоген характеризуется более разветвленной структурой, чем амилопектин, линейные отрезки цепи включают 11—18 остатков α-D-глюкопиранозы [соединенных α (1—4) -гликозидными связями], в точках ветвления остатки соединены α (1—6) -гликозидными связями (Рисунок 21).
Запасающие полисахариды бактерий
Самый распространенный полисахарид бактерий – гликоген, чья структура была рассмотрена в предыдущем разделе. Но также встречаются и другие типы (Рисунок 22).

Рисунок 22. Структура запасающих полисахаридов бактерий
Декстран. Это линейный редуцирующий гомополисахарид α D-глюкозы, мономеры соединяются 1—6 О-гликозидной связью. Декстран представляет собой линейную или слаборазветвленную молекулу, молекулярная масса которой составляет примерно 106. Данный полимер очень долго привлекал только ученых, но затем было найдено его практическое применение. Продукты частичного гидролиза этого полисахарида используются как добавка в плазмозаменителях, для создания естественного онкотического давления, что сильно улучает свойства искусственных заменителей плазмы при восполнении кровопотерь.
Леван. Запасающий полисахарид Bacillus subtilis. Линейный нередуцирующий гомополисахарид, образованный остатками метилового эфира β D фруктозы, мономеры соединяются 1—6 гликозидной связью. Полимер отличается небольшой длиной 10—12 мономеров.
Запасающие полисахариды растений
Крахмал. Моносахаридные остатки соединены в крахмале α-гликозидными связями (Рисунок 23). Соединение такой структуры, образованное только остатками глюкозы, является гомополимером, его называют глюкозаном или глюканом. Это наиболее важный вид пищевых углеводов; он содержится в злаках, картофеле, бобовых и в других растениях. Двумя главными компонентами крахмала являются амилоза (15—20%), имеющая неразветвленную спиральную структуру, и амилопектин (80—85%), образованный разветвленными цепями, каждая ветвь состоит из 24—30 остатков глюкозы, соединенных (1—4) – связями [в точках ветвления остатки соединены (1—6) – связями]. Благодаря геометрическим особенностям α (1—4) – связей линейные участки полимерных цепей в молекулах гликогена и крахмала стремятся принять скрученную, спиральную конформацию, что способствует образованию плотных гранул, которые и обнаруживаются в большинстве растительных клеток. α (1—4) -связи гликогена и крахмала легко гидролизуются α-амилазой желудочно-кишечного тракта позвоночных, а образующаяся при этом D-глюкоза попадает в кровь и далее используется в энергетическом обмене. Крахмал как и гликоген является редуцирующим полисахаридом.
Декстринами называют вещества, образующиеся при частичном гидролизе крахмала. Название «остаточные декстрины» получили продукты, образующиеся на определенной стадии гидролиза.
Фруктаны. Это линейные, редуцирующие гомополимеры β фруктозы, соединенной β (2—6) гликозидной связью (Рисунок 23). Фруктаны встречаются у различных видов растений: ирисисин в корневищах ириса, аспарогозин в корнях спаржи, секалин – в ржи. Различия фруктанов, выделенных из различных объектов, связаны с вариабельностью молекулярной массы и количества мономеров, входящих в состав полисахарида.

Рисунок 23. Структурные и запасающие полисахариды растений. А-крахмал; Б-схема организации молекулы крахмала; В-инулин; Г-фруктаны; Д-целлюлоза; Е-ксилоза; Ж– пектины
Инулин – линейный нередуцирующий полисахарид, содержащийся в клубнях и корнях георгинов, артишоков и одуванчиков. При его гидролизе образуется фруктоза, следовательно он представляет собой фруктозан. Гетерополимер одной молекулы глюкозы соединенной с β фруктозой β (1—1) гликозидной связью, остальные мономеры β фруктозы, соединяются β (1—2) гликозидной связью (Рисунок 23). Этот полисахарид в отличие от картофельного крахмала легко растворяется в теплой воде; его используют в физиологических исследованиях для определения скорости клубочковой фильтрации в почках. Также этот полимер, состоящий в основном из фруктозы, привлекает диетологов и врачей как заменитель крахмала для больных сахарным диабетом. У этих больных нарушено поглощение глюкозы клетками, тогда как фруктоза поглощается клетками больных, поэтому может быть использована как замена глюкозе. Эта же причина вызвала большой интерес к растениям запасающим фруктаны. И инулин и другие полимеры фруктозы могут использованы в диете больных сахарным диабетом, как заменители крахмала.
Структурные полисахариды растений
Целлюлоза является линейным, редуцирующим неразветвленным гомополисахаридом, состоящим из 10000 и более остатков D-глюкозы, связанных друг с другом (1—) – гликозидными связями; в этом отношении она сходна с амилозой и линейными участками цепей гликогена. Но между этими полисахаридами существует одно очень важное различие: в целлюлозе (1—4) -связи имеют β-конфигурацию, а в амилозе, амилопектине и гликогене – α-конфигурацию.
Это, казалось бы, незначительное различие в строении целлюлозы и амилозы приводит к весьма существенным различиям в их свойствах. Что касается целлюлозы, то из-за конфигурации связей ее полимерные цепи сильно вытянуты и соединяются друг с другом бок о бок, образуя длинные нерастворимые фибриллы.
Целлюлоза – прочное, волокнистое, водонерастворимое вещество – содержится в стенках клеток растений, главным образом в ветвях, стеблях, а также в стволах и других деревянистых частях растений (Рисунок 23, Д). Древесина состоит в основном из целлюлозы и других полимерных веществ, хлопок – почти целиком из целлюлозы. Если наиболее распространенные внутриклеточные биополимеры – это белки, то целлюлоза, бесспорно, это не только самый распространенный внеклеточный структурный полисахарид в растительном мире, но и вообще самый распространенный в природе биополимер.
Ежегодно огромные количества целлюлозы синтезируются растениями, причем не только растущими в лесах деревьями, но и культурными растениями. Расчеты показывают, что на долю каждого живущего на Земле человека растения ежедневно нарабатывают приблизительно 50 кг целлюлозы. Целлюлоза находит широкое применение в промышленности. Целлюлоза нерастворима в воде, кислотах, и только в концетрированных щелочах при нагревании переходит в полужидкое состояние, что используется при производстве вискозного волокна.
Связи β (1—4) в молекуле целлюлозы не гидролизуются α -амилазами. Поскольку в кишечнике позвоночных нет фермента, способного гидролизовать целлюлозу, она не переваривается, и ее D-глюкозные остатки не могут служить пищей для большинства высших организмов.
Целлюлозу хорошо переваривают термиты, но лишь потому, что в их кишечнике живут паразитические микроорганизмы Trichonympha, секретирующие целлюлазу, гидролизующий целлюлозу фермент, с помощью которого и происходит переваривание древесины у термитов. Целлюлазу синтезируют также некоторые бактерии и грибы, вызывающие гниение древесины.
Среди позвоночных только крупный рогатый скот и другие жвачные (овцы, козы, верблюды, жирафы и т. д.) могут использовать целлюлозу в качестве пищи, однако делают они это весьма необычным способом: большая часть кишечника, составляющая 15% общего веса коровы, приходится на долю четырех последовательно соединенных друг с другом желудков. Первые два из них составляют так называемый рубец. Содержащиеся в нем микроорганизмы секретируют целлюлазу и расщепляют целлюлозу до D-глюкозы, которую далее сбраживают до короткоцепочечных жирных кислот, двуокиси углерода и газообразного метана (СН4). Образовавшиеся жирные кислоты всасываются в кровоток коровы, проникают в ткани и используются как топливо. Метан и СО2, которые вырабатываются со скоростью 2 л/мин, постоянно выводятся посредством непроизвольного процесса, напоминающего едва уловимую на слух отрыжку. В остальных двух желудках жвачных микроорганизмы, сделавшие свое дело, перевариваются ферментами, секретируемыми слизистой желудка; при этом образуются аминокислоты, сахара и другие продукты, которые всасываются и используются в организме коровы в качестве питательных веществ.
Таким образом, между коровой и населяющими ее рубец микроорганизмами устанавливаются отношения симбиоза, при котором микроорганизмы получают возможность насладиться короткой, но счастливой жизнью в удобной и теплой среде; при этом целлюлоза из клевера и другой травы служит основным источником топлива и для «жильцов», и для организма-хозяина.
Гемицеллюлозы
Это многообразные полисахариды растений входящих в клеточную стенку, примерами являются ксилоза и пектины.
Ксилоза. Линейный редуцирующий гомополимер ксилозы, в β-пиранозной форме, мономеры соединяются β (1—4) гликозидной связью (Рисунок 23, Е).
Пектины. Эти полимеры встречаются очень часто в клеточных стенках плодов и отвечают за их «желирующие» свойства (чем больше пектинов, тем плотнее джемы и повидло). Это линейный редуцирующий гомополимер, мономером является метиловый эфир галактуроновой кислоты (Рисунок 23, Ж). Мономеры соединяются α (1—4) гликозидной связью. Пектиновыми веществами богаты плоды айвы, некоторых сортов груш и яблок, они обеспечивают процесс «желирования», то есть образования более плотного мармелада. Это свойство до последнего времени привлекало только кондитеров. Но в последнее время этим молекулам уделяется большое внимание, так как было показано, что пектины участвуют в выводе солей тяжелых металлов и радионуклидов. Поэтому пектины и продукты их частичного гидролиза используются как антирадиационные препараты или добавки к ним.
Структура клеточных стенок растений
Клеточная стенка растений формируется при участии плазматической мембраны и является экстраклеточным (внеклеточным) многослойным образованием, защищающим поверхность клетки и служащим, как бы наружным скелетом растительной клетки. Клеточная стенка растений состоит из двух компонентов: аморфного пластичного гелеобразного матрикса (основы) с высоким содержанием воды и опорной фибриллярной системы (Рисунок 24). Эта структура напоминает железобетон. Роль железной арматуры выполняют фибриллы из пучков молекул целлюлозы, образующие трехмерную сеть. Пространство между молекулами «арматуры» заполняют гемицеллюлозы, образующие желеобразный матрикс.
Дополнительные полимерные вещества и соли, часто входящие в состав оболочек, придают им жесткость и делают их несмачиваемыми. В химическом отношении главные компоненты оболочек растений относятся к структурным полисахаридам. В состав матрикса оболочек растений входят гетерогенные группы полисахаридов, растворяющиеся в концентрированных щелочах, гемицеллюлозы и пектиновые вещества. Гемицеллюлозы представляют собой ветвящиеся полимерные цепи, состоящие из различных гексоз (глюкоза, манноза, галактоза и др.), пентоз (ксилоза, арабиноза) и урановых кислот (глюкуроновая и галактуроновая). Эти компоненты гемицеллюлоз сочетаются между собой в разных количественных отношениях и образуют разнообразные комбинации.
Цепи гемицеллюлозных молекул не кристаллизуются и не образуют элементарных фибрилл, они образуют аморфное гелеобразное вещество. Из-за наличия полярных групп урановых кислот они сильно гидратированы. Пектиновые вещества – гетерогенная группа, в которую входят разветвленные, сильно гидратированные полимеры, несущие отрицательные заряды из-за множества остатков галактурононой кислоты. Благодаря свойствам своих компонентов матрикс представляет собой мягкую пластическую массу, укрепленную фибриллами. Волокнистые компоненты клеточных оболочек растений состоят обычно из целлюлозы – линейного, неветвящегося полимера глюкозы. Молекулярная масса целлюлозы варьирует от 5—104 до 5—105, что соответствует 300—3000 остаткам глюкозы. Такие линейные молекулы целлюлозы могут соединяться в пучки или волокна.

Рисунок 24. Схема строения клеточной стенки растений; 1 – первичная оболочка (два слоя по обе стороны от 2); 2 – срединная пластинка; 3 – слои вторичной оболочки; 4—третичная оболочка
В клеточной оболочке целлюлоза образует фибриллы, которые состоят из субмикроскопических микрофибрилл толщиной до 25 нм, а они в свою очередь состоят из множества параллельно лежащих цепей молекул целлюлозы.
Количественные соотношения целлюлозы к веществам матрикса (гемицеллюлозы) могут быть весьма различными у разных объектов. Свыше 60% сухой массы первичных оболочек составляет их матрикс и около 30% приходится на скелетное вещество – целлюлозу. В сырых клеточных оболочках почти вся вода связана с гемицеллюлозами, поэтому масса основного вещества в набухшем состоянии достигает 80% сырой массы всей оболочки, тогда как содержание волокнистых веществ сводится всего к 12%. В волосках хлопчатника целлюлозный компонент составляет 90% в древесине на долю целлюлозы приходится 50% от компонентов клеточной стенки.
Кроме целлюлозы, гемицеллюлозы и пектинов в состав клеточных оболочек входят дополнительные компоненты, придающие им особые свойства. Так, инкрустация (включение внутрь) оболочек лигнином (полимер кониферилового спирта) приводит к одревеснению клеточных стенок, повышению их прочности. Лигнин замещает в таких оболочках пластические вещества матрикса, и играет роль основного вещества, обладающего высокой прочностью. Часто матрикс укреплен минеральными веществами (Si02, Са2СО3 и др.).
На поверхности клеточной оболочки могут скапливаться различные адкрустирующие вещества, например кутин и суберин, приводящие к опробковению клеток (Рисунок 25). В клетках эпидермиса на поверхности клеточных оболочек откладывается воск, который образует водонепроницаемый слой, препятствующий потере клеткой воды. Из-за своего пористого, рыхлого строения клеточная стенка растений проницаема в значительной степени для низкомолекулярных соединений, таких как вода, сахара и ионы. Но макромолекулы проникают через целлюлозные оболочки плохо: величина пор в оболочках, позволяющая свободную диффузию веществ, составляет всего лишь 3—5 нм.
Опыты с мечеными соединениями показали, что при росте клеточной оболочки выделение веществ, из которых она строится, происходит по всей поверхности клетки. Аморфные вещества матрикса, гемицеллюлозы и пектины синтезируются в вакуолях аппарата Гольджи и выделяются через плазмалемму путем экзоцитоза. Фибриллы целлюлозы синтезируются специальными ферментами, встроенными в плазмалемму. Оболочки дифференцированных, зрелых, клеток обычно многослойные, в слоях фибриллы целлюлозы ориентированы по-разному, и количество их также может значительно колебаться. Обычно описывают первичные, вторичные и третичные клеточные оболочки. Для того чтобы разобраться в строении и появлении этих оболочек, необходимо познакомиться с тем, как они образуются после деления клеток.

Рисунок 25. Инкрустация клеточной оболочки. А – фибриллярный каркас и межфибриллярный матрикс; Б – инкрустированная лигнином и утерявшая способность к растяжению оболочка с остатками матрикса; В – последующее инкрустирование фенолами и (или) минеральными веществами, приводящее к повышению твердости оболочки
Структурные полисахариды грибов

Рисунок 26. Структурные полисахариды грибов
Клеточные стенки грибов с одной стороны плохо изучены по структуре, а с другой очень разнообразны. Именно поэтому можно говорить о полисахаридах, составляющих основу клеточной стенки грибов, но не о структуре клеточной стенки.
У различных групп грибов клеточная стенка формируется несколькими типами полисахаридов. У оомицетов (к ним относится возбудитель фитофторы) клеточная стенка образуется целлюлозой. У дрожжей – маннаны. Маннаны – редуцирующие гомополимеры β-D-маннозы, моносахариды соединяются β (1—6) гликозидной связью. Полимер разветвленный. «Ветки» присоединяются β (1—2) связью, такой же связью соединены мономеры в составе ответвления. В результате образуется сложный пересекающийся каркас, по структуре напоминающий войлок. Дополнительное укрепление связи молекул обеспечивают белки клеточной стенки. У базидиомицетов клеточную стенку формирует хитин (Рисунок 26). Длинный неразветвленный редуцирующий гомополисахарид. Структуру хитина составляют N N-ацетил-О-глюкозаминовые звенья, соединенные β (1—4) -гликозидными связями.
Структурные полисахариды бактерий
Клеточная стенка бактерий образует вокруг клетки жесткую пористую оболочку. Она физически защищает нежную клеточную мембрану и цитоплазму клетки. Структурной основой клеточных стенок большинства бактерий служит пронизанный поперечными ковалентными связями каркас, который почти целиком окружает клетку (Рисунок 27).

Рисунок 27. Формулы структурных полисахаридов бактерий
Он состоит из длинных, параллельно расположенных полисахаридных цепей, связанных между собой через определенные интервалы поперечными сшивками из коротких полипептидных цепочек. Полисахаридные цепи состоят из чередующихся моносахаридных остатков N-ацетил-D-глюкозамина и N-ацетилмурамовой кислоты (сложного девятиатомного сахара), соединенных друг с другом β (1 -4) -связями. К каждому остатку N-ацетилмурамовой кислоты присоединена боковая тетрапептидная цепочка, образованная последовательно соединенными L-аланином, D-глутаминовой кислотой, мезо-диаминопимелиновой кислотой, D-аланином. В результате параллельные полисахаридные цепи сшиваются короткими поперечными полипептидными цепочками, структура которых различна у разных видов бактерий. У гноеродных бактерий Staphylococcus aureus, вызывающих развитие фурункулов и нагноение ран, остатки ацетилмурамовой кислоты в соседних полисахаридных цепях связаны друг с другом пептидными цепочками, состоящими из пяти остатков глицина.
Вся эта скрепленная поперечными связями структура, окружающая клетку, называется муреином или пептидогликаном; второе название подчеркивает гибридную природу данной структуры, представляющей собой сочетание пептидных и полисахаридных элементов. Тянущийся непрерывно вдоль всей поверхности бактериальной клетки пептидогликан можно рассматривать как одну гигантскую мешковидную молекулу, по структуре похожую на сетку. У грамположительных бактерий (дающих окраску по Граму, т. е. при обработке красителем кристаллическим фиолетовым) пептидогликан образует вокруг клетки около 60 концентрических слоев, пронизываемых другими макромолекулярными компонентами, прежде всего тейхоевыми кислотами.
У грамотрицательных бактерий, например у Е. coli, пептидогликановый каркас покрыт богатой липидами внешней оболочкой, содержащей гидрофобные белки. Целостность клеточных стенок имеет жизненно важное значение для защиты, роста и деления бактерий. Действие пенициллина – одного из наиболее ценных антибиотиков, используемых для борьбы с бактериальными инфекциями, основано на том, что он подавляет последний этап ферментативного синтеза пептидогликанов у чувствительных к нему микроорганизмов; это приводит к формированию неполноценных клеточных стенок и подавлению роста бактерий.
Структура клеточных стенок бактерий

Рисунок 28. Структура клеточных стенок бактерий. А – грамположительных; Б – грамотрицательных. В – Структурная формула глицеринтейхоевой кислоты. Содержит чередующиеся остатки D-аланина и N-ацетилглюкозамина
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотических организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы (Рисунок 28).
Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом, в зависимости от строения клеточной стенки, судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу, так и по ультраструктуре. В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке.
У грамположительных эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90%), у грамотрицательных – содержание пептидогликана значительно меньше (1—10%). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50% этого гетерополимера. Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм.
У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2—3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8—10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны. Клеточная стенка грамположительных эубактерий плотно прилегает к мембране в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от мембране.
Пространство между цитоплазматической и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм. Клеточная стенка грамположительных эубактерий.
Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер – пептидогликан. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы. Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы – в формировании дополнительных пептидных связей между гетерополимерными цепочками. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободная аминогруппа диаминопимелиновой кислоты другого. Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот. Нетрудно себе представить, что этим способом можно «сшить» между собой множество гетерополимерных цепей. Частота «сшивок» различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии.
Пептидогликан, окружающий протопласт грамположительных эубактерий, – это по существу одна гигантская молекула, «сшитая» с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей трехмерную пространственную организацию. Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений – тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями. Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с N-ацетилмурамовой кислотой. Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных эубактерий. Остающиеся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона. Как полианионы тейхоевые кислоты определяют поверхностный заряд клетки. Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фага на клеточной поверхности.
В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки и липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой.
Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией. Клеточная стенка бацилл, например Bacillus subtilis, приблизительно соответствует толщине 40 молекул пептидогликана.
В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1 – 6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор.
Клеточная стенка грамотрицателъных эубактерий.
У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных. В ее состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к мембране. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями.
Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки – наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида. Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30 – 40% ее поверхности и локализованный во внешнем слое.
Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80% всех белков наружной мембраны. Одна из функций этих белков – формирование в мембране гидрофильных пор диаметром примерно 1 нм, через которые осуществляется неспецифическая диффузия молекул с массой до 600 – 900 Да. Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды.
Структурные полисахариды животных
Хитин – важный структурный полисахарид беспозвоночных. Из него, в частности, построен наружный скелет ракообразных и насекомых. Структуру хитина составляют N N-ацетил-О-глюкозаминовые звенья, соединенные β (1—4) -гликозидными связями.
Гликозаминогликаны (мукополисахариды) У позвоночных входят в состав межклеточного вещества в составе протеогликанов (соединения из углеводов и белков где углевод составляет 90%).

Рисунок 29. Формулы структурных полисахаридов позвоночных. А-гиалуроновая кислота, Б-гепарин, В-хондроэтин-4-сульфат
Гиалуроновая кислота образует очень вязкие, гелеобразные растворы, входит в состав рыхлой и плотной соединительных тканей, а также в состав хряща. состоящий из многократно чередующихся остатков D-глюкуроновой кислоты и N-ацетил-О- глюкозамина, соединенных β (1—3) гликозидной связью. Это линейный редуцирующий гетерополисахарид.

Рисунок 30. Структура протеогликанов
Хондроитин, основной полисахарид протеогликанов хряща, это линейный, редуцирующий гетерополисахарид, содержащий чередующиеся остатки D-глюкуроновой кислоты и сульфатированного N-ацетил-D-галактозамина, соединенные β (1—3) гликозидной связью. В зависимости от сульфатирования, есть 4- или 6-хондроитинсульфат. Также в состав межклеточного вещества соединительной ткани входят кератансульфат и другие полисахариды (Рисунок 30). Существует еще множество полисахаридов, входящих в состав протеогликанов, образующих аморфное вещество соединительных тканей. В зависимости от типа соединительной ткани преобладает тот или иной тип полисахарида в протеогликанах. Все они слабо растворимы, но существует исключение – полисахарид плазмы крови, синтезируемый клетками, выстилающими сосуды. Этот полисахарид – гепарин.
Гепарин – это короткий линейный, редуцирующий гетерополисахарид, растворенный в плазме крови. В состав гепарина входят повторяющиеся единицы из остатков шести сахаров, каждая из которых представляет собой последовательность чередующихся остатков сульфопроизводных N-ацетил – D-глюкозамина и D-идуроната. Гепарин препятствует свертыванию крови, то есть является антикоагулянтом. Его секретируют клетки выстилающие капилляры – эндотелиоциты. Выделенный из легочной ткани гепарин используется в медицине для предотвращения свертывания донорской крови, а также для предупреждения свертывания крови в сосудах при различных патологических состояниях, например, после приступов стенокардии.
Структурные полисахариды, входящие в состав межклеточного вещества, образуют протеогликаны – белково-углеводные комплексы, где белок составляет около 10%. Типичный протеогликан хрящевой ткани содержит около 150 полисахаридных цепей с молекулярной массой 20000 каждая; они (в виде боковых цепей) ковалентно присоединены к «сердцевинным» полипептидам. Такие протеогликаны представляют собой сильно гидратированные структуры, 20000 каждая; они (в виде боковых цепей) ковалентно присоединены к «сердцевинным» полипептидам. Такие протеогликаны представляют собой сильно гидратированные структуры.
Гликопротеиды – это молекулы белка, к которым присоединены короткие полисахаридные молекулы. В отличие от протеогликанов, в гликопротеидах белок составляет 95% молекулы. Кроме того, углеводный компонент короче – несколько десятков мономерных звеньев, очень разнообразен, можно сказать, что каждый белок имеет свой уникальный как по составу, и по форме полисахарид. Полисахариды присоединяются к секретируемым белкам клетки, а также к мембранным белкам, углеводная часть протеидов мембраны обращена во внешнюю среду клетки, формирует гликокаликс.
В данном случае полисахарид выполняет сигнальную функцию, по таким разветвленным гетерополисахаридам клетки распознают друг друга. В частности группы крови А В определяются по полисахаридам располагающимся на поверхности эритроцитов. Здесь можно говорить не только о многообразии полисахаридного компонента каждого белка, но и о вариабельности этой молекулы для одного белка внутри популяции.