Глава 1. Происхождение жизни
Вопрос о происхождении жизни волнует всех, и очень жаль, что он пока еще далек от разрешения. Основная сложность тут в том, что путь от неорганических молекул к первой живой клетке был долгим и трудным. За один шаг такие превращения не происходят (если, конечно, не привлекать сверхъестественные силы). На этом долгом пути было множество развилок, остановок, временных отступлений, всевозможных промежуточных этапов и "переходных звеньев". Досаднее всего, что эти многочисленные, разнообразные и сложные события не оставили, по-видимому, никаких следов в палеонтологической летописи. Земная кора просто не сохранила столь древних осадочных пород. К тому же есть основания полагать, что какие-то ранние этапы пути могли быть пройдены и вовсе не на Земле, а в протопланетном облаке или на других небесных телах. Что касается "переходных звеньев", то все они благополучно вымерли, не оставив после себя прямых потомков, за исключением одного-единственного – так называемого "последнего универсального общего предка" всех современных живых существ. Специалисты называют его Лукой (Last Universal Common Ancestor, LUCА). Лука был уже довольно сложным организмом, напоминающим бактерию.
Поэтому ученые пока вынуждены довольствоваться разработкой правдоподобных гипотез, описывающих и объясняющих отдельные этапы долгого пути от мертвой материи к живой клетке. Эти гипотезы поддаются экспериментальной проверке, но таким способом можно доказать только принципиальную возможность того или иного хода событий. Гораздо труднее доказать, что все на самом деле именно так и происходило.
Тем не менее даже в этой крайне трудной области исследователи в последние годы добились заметных успехов. Начать, однако, лучше с истории.
Маленькая колба с бульоном порождает большие проблемы
Наши далекие предки и не подозревали, что вопрос о происхождении живого из неживого может стать серьезной проблемой для их потомков. Для них все вокруг было живым, одухотворенным: солнце и воздух, горы и реки, облака и море. Мудрецы Античности и Средневековья тоже не видели непреодолимой грани между живым и неживым. Вслед за Аристотелем вплоть до xvii века все ученые считали зарождение жизни самым обычным, повседневным явлением. В гниющих отходах зарождаются черви и мухи, в старом тряпье – мыши, на подводных камнях и днищах кораблей – моллюски. Могучая "животворная сила" пронизывает мироздание; она-то и заставляет косную материю порождать жизнь. Это учение – витализм – не противоречило и библейской версии космогенеза.
"И сказал Бог: да произрастит земля зелень…"
"И сказал Бог: да произведет вода пресмыкающихся…"
Бог дал стихиям творческие силы. И с тех пор – чему же удивляться? – вода производила, а земля произращивала.
Первым усомнился итальянец Франческо Реди (1626–1698), заявивший, что всякий живой организм происходит только от другого живого организма. Реди доказал, что личинки мух не заведутся в тухлом мясе, если мясо накрыть тряпочкой, сделав его недоступным для взрослых мух. Публика согласилась с Реди, что такие сложные животные, как мухи и мыши, могут рождаться только от соответствующих родителей. Но вот простые одноклеточные организмы – они-то, несомненно, порождаются грязной водой, гниющими отходами и тому подобным. Эта концепция просуществовала в общественном сознании около ста лет, а затем ученый монах Ладзаро Спалланцани, воодушевленный открытиями Левенгука, обнаружил, что микробы тоже размножаются – делятся пополам, давая начало таким же микроорганизмам. Мало того, когда он кипятил бульон и плотно закрывал его крышкой, никаких микробов там не появлялось.
Конечно, это был чисто научный спор о зарождении живых существ, так сказать, исследование начала начал, но и кое-какая практическая польза из этого вышла. Об опытах Спалланцани прознал французский повар по имени Николя Аппер. В микробах, происхождении жизни, витализме и прочих энтелехиях он ничего не понимал, зато отлично разбирался в мясных подливках и бульонах, которые Спалланцани использовал в качестве питательной среды. А как раз в это время Наполеон ввиду грядущих военных кампаний был обеспокоен вопросами снабжения армии. Ведь армейская кухня вынуждена была вести за собой в обозе целые стада и грабить население – а это для дальновидного военачальника ненадежные и неудобные способы снабжения. Поэтому Наполеон в 1804 г. подал в Сенат прошение об учреждении премии тому, кто изобретет способ долгого хранения продуктов. Сенат, естественно, так и сделал (Наполеон к этому времени уже был пожизненным Первым консулом), учредив внушительную по тем временам премию в 12 тыс. франков. И Аппер решил во что бы то ни стало ее получить. Повар подумал, что ученый монах подкинул неплохую идею: сделать жаркое, прокипятить его, положить в чистую банку и хорошенько закупорить. Так были изобретены консервы, и изобретателем их по сей день считается Аппер. А нам с вами нужно хорошо понимать, что люди с практической сметкой даже из самого отвлеченного теоретического знания могут извлечь пользу – если, конечно, эти теории покоятся на реальных законах природы.
Но вернемся от прикладных применений к теоретическим вопросам происхождения жизни, какими они представлялись после открытий Спалланцани. Стало ясно, что микробы, как и обычные живые существа, размножаются, дают потомство, а вовсе не самозарождаются. Однако многие ученые считали, что для зарождения жизни необходима "жизненная сила". Она не может проникнуть в закрытую банку, именно потому там и не появляются микробы. Как это проверить? Жизненную силу приборами не измерить и руками не ухватить. Луи Пастер в 1865 году хитроумным экспериментом сумел доказать, что никакой жизненной силы нет. Этот опыт, показанный в парижской Академии наук во время знаменитого диспута с Пуше, вошел в школьные учебники всех стран. Пастер предложил не запаивать наглухо прокипяченный бульон, а оставить в стеклянной колбе тонкий, дважды изогнутый открытый носик. Если жизненная сила есть, то даже узкий извилистый проход не помешает ей проникнуть в колбу и породить микробов. Если же такой силы нет, то микробы из воздуха будут оседать на изгибах стеклянного носика, и бульон останется стерильным. Оппонент Пастера Феликс Пуше утверждал, что в колбе с тонким изогнутым носиком в питательном бульоне микробы все равно появятся. И это означало бы присутствие жизненной силы.
Пуше на тот эпохальный диспут не явился. И академики, разумеется, признали поражение Пуше и победу Пастера. Ведь если один из спорщиков не пришел на суд научной общественности, значит, он сам признал свою неправоту. Таким образом, ученый мир удостоверился, что живое порождается только живым.
Результат диспута удовлетворил всех, кроме… самого Пастера. Дело в том, что в хитроумных пастеровских колбах, как ни кипяти питательную среду, какими узкими ни делай изогнутые носики, микробы все-таки появлялись. Не должны были появляться, неоткуда им было взяться, но появлялись, и сам Пастер об этом знал. Хотя появление микроорганизмов в кипяченом бульоне противоречило его обширному опыту и здравому смыслу. Во время диспута Пастер не признался в своих сомнениях, но в течение следующих го лет пытался эту загадку разрешить. И разрешил. Оказалось, что дело тут в специфике того микроба, с которым работал Пуше. Это была сенная палочка, споры которой погибают только при температуре 120 °C. Кипячением эти споры не уничтожаются, и для того, чтобы все же доказать самому себе победу в том давнем диспуте, Пастеру пришлось изобрести автоклав, аппарат для стерилизации при больших давлениях и температурах. Так что, можно сказать, результатом того исторического спора оказалось не только доказательство отсутствия жизненной силы, но и изобретение автоклава. О пользе первого можно спорить, а вот автоклавом люди пользуются до сих пор.
Неотъемлемое свойство науки – самой себе создавать проблемы. С витализмом было покончено, но все дивиденды достались не науке, а… церкви. Живое не может самозарождаться? Превосходно! Но откуда тогда вообще появилась жизнь? Одно из двух: либо она существовала изначально (эту точку зрения позже отстаивал В.И. Вернадский), либо сотворена Богом (так полагал и сам Луи Пастер). Второе, конечно, вероятнее: представить себе "изначальность" жизни (как и вечность Вселенной) довольно трудно. А вот теория божьего творения проста и понятна каждому.
Нелегкая задача встала перед теми учеными, которые не соглашались с Луи Пастером и не хотели привлекать сверхъестественные силы для объяснения природных явлений. После того как была доказана невозможность самозарождения, они должны были теперь долго и мучительно доказывать его возможность. Пусть не здесь и сейчас, а очень давно, и не за час-другой, а за миллионы лет.
Поначалу дело казалось безнадежным, ведь химики еще не умели получать органические вещества из неорганических. Грань между живой и неживой материей казалась непреодолимой. Правда, еще в 1828 г. Фридрих Велер синтезировал мочевину, но это никого не убедило: подумаешь, мочевина! Не органика, а так, "отход жизнедеятельности". Однако в 1854 г. Пьер Эжен Марселей Бертло сумел искусственно получить липиды, а в 1864 г. А. М. Бутлеров открыл реакцию синтеза углеводов из формальдегида. Вскоре химики научились получать и многие другие органические вещества из неорганических.
Стало ясно, что между живой и неживой материей на химическом уровне нет непреодолимой грани. Это был первый крупный успех теории абиогенеза (происхождения живого из неживого естественным путем). Ученые поняли, что, хотя прямое самозарождение живых существ невозможно, жизнь могла появиться постепенно в результате долгой химической эволюции. С тех пор (и до сего дня) усилия ученых направлены на поиски доказательств и развитие этой гипотезы. Что касается идеи об изначальности жизни, то она сейчас практически не имеет сторонников, поскольку наших знаний о Вселенной уже достаточно, чтобы понимать – на первых этапах ее развития синтез даже самых простых органических соединений был невозможен. Третий вариант – божественное сотворение – не имеет проверяемых следствий (поскольку Бог мог придать сотворенной им жизни абсолютно любые свойства, о которых мы ничего не можем знать заранее) и не предлагает никакой исследовательской программы. Поэтому он не является научным и просто-напросто не интересен ученым. Тем не менее ученым пришлось бы отнестись к нему более серьезно, если бы были получены строгие доказательства несостоятельности теории абиогенеза. Эта теория, в отличие от креационизма, является научной, то есть допускающей принципиальную возможность опровержения. Например, если бы оказалось, что органические вещества действительно невозможно получить из неорганических, как думали многие вплоть до опытов Бертло и Бутлерова, то абиогенез пришлось бы признать несостоятельной теорией. Однако этого не происходит. Наоборот, по мере развития науки один за другим проясняются все новые этапы долгого пути от неживой природы к первой клетке.
Рождение Вселенной: первые подступы к жизни
В последние годы развитие Вселенной часто описывают как единый направленный процесс, в ходе которого самопроизвольно и неизбежно возникали все более сложные структуры[9]. Возникло особое междисциплинарное направление исследований – Универсальная История, или Big History. Один из его основоположников – профессор Дэвид Кристиан из университета Сан-Диего, между прочим, специалист по истории России. При взгляде на историю мироздания "с высоты птичьего полета" создается впечатление, что каждый новый шаг в эволюции Вселенной логически вытекал из предыдущего и в свою очередь предопределял следующий. Возникновение жизни предстает уже не случайностью, а закономерным итогом развития. Вселенная словно была изначально спроектирована так, чтобы в ней появилась жизнь, и проект был чрезвычайно точен. Даже небольшое изменение базовых физических констант сделало бы жизнь невозможной (по крайней мере такую жизнь, как наша). Впрочем, в такой Вселенной некому было бы и рассуждать о мудрости ее устройства. Кто знает, может быть, существует много разных Вселенных, и только в нашей все так удачно сложилось? Физики относятся к такой возможности вполне серьезно. И потом, развитие жизни идет по пути приспособления, адаптации, так что вполне естественно, что жизнь в нашей Вселенной соответствует ее законам. В другом мироздании и жизнь была бы другой. С этой точки зрения соответствие земной жизни земным условиям не более удивительно, чем соответствие формы воды, налитой в сосуд, форме сосуда.
Физики утверждают, что Вселенная появилась в результате Большого взрыва из очень малого и очень плотного объекта примерно 14 миллиардов лет назад (цифра пока не окончательная, она продолжает уточняться). В первые мгновения не было даже атомов и молекул. Вселенная стремительно расширялась и остывала. В определенный момент появились элементарные частицы, из них образовались атомы водорода; скопления атомов превратились в звезды первого поколения. В них происходила реакция ядерного синтеза, в ходе которой водород превращался в гелий, а затем и в более тяжелые элементы. Все необходимые для жизни элементы, кроме водорода (углерод, кислород, азот, фосфор, сера и другие), образовались в недрах звезд. Звезды первого поколения стали фабрикой по производству атомов, необходимых для будущей жизни.
Самые крупные звезды после истощения запасов ядерного топлива взрывались (это называют "взрывом сверхновой"). В результате таких взрывов атомы тяжелых элементов рассеивались в пространстве. Из новых скоплений атомов образовались звезды второго поколения, в том числе и наше Солнце. Облака рассеянных частиц, не вошедших в состав центральной звезды, вращались вокруг нее и постепенно разделялись на отдельные сгустки – будущие планеты. Именно на этом этапе и мог начаться синтез первых органических молекул.
Возможность органического синтеза в протопланетном облаке предполагалась давно, но подтверждена была лишь недавно, во многом благодаря работам академика В. Н. Пармона и его коллег из Новосибирского института катализа. При помощи сложных расчетов и компьютерного моделирования было показано, что в газово-пылевых протопланетных облаках имеются необходимые условия для синтеза разнообразной органики из водорода, азота, угарного газа, цианистого водорода и других простых молекул, вполне обычных в космосе. Непременным условием является присутствие твердых частиц-катализаторов, содержащих железо, никель и кремний[10].
Молодая Земля могла иметь в своем составе большое количество органики с самого начала своего существования. Абиогенный синтез органики продолжался уже на Земле. Экспериментально показана возможность синтеза аминокислот и других органических соединений в тех условиях, которые предположительно могли существовать на планете в дни ее молодости[11].
Гидротермальные источники – колыбель жизни на Земле?
Недавно немецким химикам удалось доказать возможность абиогенного синтеза органических веществ в условиях, которые и по сей день существуют на дне океанов. Оказалось, что в подводных горячих вулканических источниках могут происходить химические реакции, в результате которых из неорганических соединений, таких как угарный газ (СО) и цианистый водород (HCN), образуются разнообразные органические молекулы. Катализатором этих реакций служат присутствующие в гидротермальных водах твердые частицы, содержащие железо и никель. Реакции особенно хорошо идут при температуре 80-120 градусов. Условия, в которых проводились эксперименты, были максимально приближены к реальности. По мнению исследователей, такие условия (включая все компоненты реакционной смеси) вполне могли существовать в подводных вулканических источниках на ранних этапах развития Земли.
Основным продуктом реакций были гидроксикислоты и аминокислоты. В небольших количествах были получены и другие органические вещества, в том числе и такие, из которых в несколько иных условиях могут синтезироваться сахара и липиды. В отличие от других известных опытов по абиогенному синтезу органики, где не было железо-никелевых катализаторов и применялись "ударные" воздействия вроде электрических разрядов, в искусственно воссозданных условиях гидротермальных источников реакции протекали без образования "отходов" – инертных углеводородных смесей вроде дегтя или смол, которые в дальнейшем очень трудно превратить в вещества, необходимые для самозарождения жизни.
Открытие немецких химиков – весомый аргумент в пользу гипотезы, согласно которой жизнь на Земле могла зародиться в подводных вулканических источниках.
Преджизнь
Вместе с Землей возник и так называемый "геохимический круговорот". Одни вещества поступали из сдавленных, разогревшихся недр Земли, формируя первичную атмосферу и океаны. Другие приходили из космоса в виде падающих с неба остатков протопланетного облака, метеоритов и комет. В атмосфере, на поверхности суши и в водоемах все эти вещества смешивались, вступая друг с другом в химические реакции, и превращались в новые соединения, которые в свою очередь тоже вступали в реакции друг с другом.
Между химическими реакциями возникала своеобразная конкуренция – борьба за одни и те же субстраты (исходные вещества, необходимые для их проведения). В такой борьбе всегда побеждает та реакция, которая идет быстрее. Так что среди химических процессов начинается настоящий естественный отбор. Медленные реакции постепенно затухают и прекращаются, вытесняемые более быстрыми.
Важнейшую роль в этом соревновании играли катализаторы – вещества, ускоряющие те или иные химические превращения. Огромное преимущество должны были получать реакции, катализируемые своими собственными продуктами. Такие реакции называют автокаталитическими или цепными. Типичный пример автокаталитической реакции – так называемая реакция Бутлерова, в ходе которой из формальдегида образуются сахара, которые сами и являются катализаторами этой реакции. Это значит, что после появления в реакционной смеси первых молекул сахаров процесс превращения формальдегида в сахара начинает самопроизвольно ускоряться и становится лавинообразным[12].
Следующий этап – формирование автокаталитических циклов, в ходе которых происходит не только синтез катализаторов, но и частичное возобновление расходуемых субстратов. От сложного и эффективного автокаталитического цикла уже недалеко и до настоящей жизни, ведь жизнь в основе своей – это самоподдерживающийся, автокаталитический процесс.
Проблема возникает с "затравкой": та же реакция Бутлерова не начнется, если в среде изначально не будет хотя бы небольшого количества углевода-катализатора. Несколько упрощает проблему гипотеза, предложенная А. Д. Пановым из Института ядерной физики. Известно, что небесные тела могут обмениваться веществом: при столкновении планеты с крупным астероидом из ее поверхности выбиваются фрагменты породы, которые могут улететь в космос и попасть на другие планеты. По расчетам Панова, благодаря такому "метеоритному обмену" возникшее в ходе химической эволюции на одной из планет полезное новшество (например, эффективный катализатор) может в течение обозримого времени попасть в другие звездные системы, а за несколько сотен миллионов лет ареал распространения новшества может охватить всю галактику – разумеется, при условии, что запас данного вещества будет возобновляться в цепочке спровоцированных им химических превращений. Гипотеза Панова расширяет масштаб химической "кухни", в которой подготавливались ингредиенты будущей жизни, от планетарного до галактического[13].
Главная молекула жизни
Общепринятого определения жизни не существует (в биологии вообще с определениями трудно – обычно чем строже определение, тем хуже оно работает). Одни ученые полагают, что жизнь – скорее процесс, чем структура, и определяют ее, например, как "процесс сохранения неравновесного состояния органической системы извлечением энергии из среды". Такому определению могут соответствовать и системы, не имеющие четких пространственных границ, – автокаталитические циклы, "живые растворы". Другие подчеркивают обязательную дискретность живых объектов и считают, что понятие "жизнь" неотделимо от понятия "организм". Третьи подчеркивают информационную природу жизни и определяют ее как способность некого фрагмента информации ("репликатора") к самокопированию с использованием ресурсов внешней среды. Под это определение подходят не только биологические вирусы, но и компьютерные и даже распространяющиеся в обществе слухи, верования и т. п[14]. Но это, пожалуй, чересчур широкий взгляд на жизнь.
Нам известна только одна жизнь – земная, и мы не знаем, какие из ее свойств являются обязательными для любой жизни вообще. Рискнем, однако, два таких свойства все-таки назвать. Это, во-первых, наличие наследственной информации, во-вторых – активное осуществление функций, направленных на самоподдержание и размножение, а также на получение энергии, необходимой для выполнения всей этой работы.
Все живое на Земле справляется с перечисленными задачами при помощи трех классов сложных органических соединений: ДНК, РНК и белков[15]. ДНК взяла на себя первую задачу – хранение наследственной информации. Белки отвечают за вторую: они выполняют все виды активных "работ". Разделение труда у них очень строгое. Белки не хранят наследственную информацию, ДНК не совершает активной работы (в биологии почти не бывает правил без исключений, в данном случае исключения тоже есть, но очень редкие и не делающие погоды).
Молекулы третьего класса веществ – РНК – служат посредниками между ДНК и белками, обеспечивая считывание наследственной информации. При помощи РНК осуществляется синтез белков в соответствии с записанными в молекуле ДНК "инструкциями". Некоторые из функций, выполняемых РНК, очень похожи на функции белков (активная работа по прочтению генетического кода и синтезу белка), другие напоминают функции ДНК (хранение и передача информации). И все это РНК делает не в одиночку, а при активном содействии со стороны белков. На первый взгляд РНК кажется "третьей лишней". В принципе нетрудно представить себе организм, в котором РНК вовсе нет, а все ее функции поделили между собой ДНК и белки. Правда, таких организмов в природе не существует.

РНК – полимер, состоящий из множества похожих "кирпичиков" – рибонуклеотидов, каждый из которых, в свою очередь, собран из трех частей. Первая из них – фосфорная кислота (фосфат), неорганическое вещество, которого довольно много в земной коре и океанах. Вторая – азотистое основание. В состав РНК входят четыре азотистых основания: А (аденин), У (урацил), Г (гуанин) и Ц (цитозин); соответственно, существует четыре вида рибонуклеотидов. Азотистые основания могли синтезироваться из неорганических молекул (таких как СО, HCN и NHp еще в протопланетном облаке. Их находят и в метеоритах. Третья – сахар рибоза – образуется в ходе автокаталитической реакции Бутлерова.
Четыре составных блока молекулы РНК – нуклеотиды аденозин (аденин + рибоза), гуанозин (гуанин + рибоза), уридин (урацил + рибоза) и цитидин (цитозин + рибоза), к каждому из которых присоединено по одному остатку фосфорной кислоты. АТФ (главная энергетическая молекула живой клетки) представляет собой аденозин с тремя фосфатами.

Какая из трех молекул появилась первой? Одни ученые говорили: конечно, белки, ведь они выполняют всю работу в живой клетке, без них жизнь невозможна. Им возражали: белки не могут хранить наследственную информацию, а без этого жизнь и подавно невозможна! Значит, первой была ДНК!
Ситуация казалась неразрешимой: ДНК ни на что не годна без белков, белки – без ДНК. Получалось, что они должны были появиться вместе, одновременно, а это трудно себе представить. Про "лишнюю" РНК в этих спорах почти забыли. Ведь она, как тогда думали, не может без посторонней помощи ни хранить информацию, ни выполнять работу.
Потом, правда, выяснилось, что у многих вирусов наследственная информация хранится в виде молекул РНК, а не ДНК. Но это посчитали курьезом, исключением. Переворот произошел в 80-х годах XX века, когда были открыты рибозимы – молекулы РНК с каталитическими свойствами. Рибозимы – это РНК, выполняющие активную работу, то есть то, что должны делать белки. Среди рибозимов были найдены и катализаторы репликации (копирования, размножения) молекул РНК – своих собственных или чужих.
В итоге РНК из "почти лишней" стала "почти главной". Оказалось, что она, и только она, может выполнять сразу обе главные жизненные задачи – и хранение информации, и активную работу. Стало ясно, что возможен полноценный живой организм, не имеющий ни белков, ни ДНК, в котором все функции выполняются только молекулами РНК. Конечно, ДНК лучше справляется с задачей хранения информации, а белки – с "работой", но это уже детали. РНК– организмы могли приобрести белки и ДНК позже, а поначалу обходиться без них.
Так появилась теория РНК-мира, согласно которой первые живые существа были РНК-организмами без белков и ДНК. А первым прообразом будущего РНК-организма мог стать автокаталитический цикл, образованный самовоспроизводящимися молекулами РНК – теми самыми рибозимами, которые способны катализировать синтез собственных копий.
Лично я считаю теорию РНК-мира одним из самых выдающихся достижений теоретической мысли в биологии за последние 30 лет, которое можно сравнить разве что с симбио-генетической теорией происхождения эукариот[16] (о которой пойдет речь в главе "Великий симбиоз"). По правде сказать, могли бы до этого додуматься и раньше. Ведь два вида рибозимов были известны еще с 60-х годов XX века, хотя их не называли тогда рибозимами. Это рибосомные РНК (рРНК), из которых сделаны молекулярные "машинки" для трансляции (синтеза белка) – рибосомы, и транспортные РНК (тРНК), которые подносят нужные аминокислоты к рибосомам в ходе трансляции. Оба вида РНК выполняют активную биохимическую работу, это самые настоящие рибозимы, но, пока их было известно только два, теория РНК-мира не была сформулирована.
Трансляция – синтез белка. Осуществляется особыми молекулярными "машинками" – рибосомами, которые состоят из нескольких больших молекул рибосомной РНК и большого числа менее крупных молекул рибосомных белков. Рибосомы синтезируют белок в соответствии с "инструкциями", записанными в молекуле матричной РНК (мРНК). Каждые три нуклеотида мРНК кодируют одну аминокислоту. Аминокислоты присоединяются к синтезируемой молекуле белка по одной. Доставка аминокислот к рибосомам осуществляется транспортными РНК (тРНК).
Теория РНК-мира, вначале чисто умозрительная, очень быстро "обрастает" экспериментальными данными. Химики научились получать рибозимы чуть ли не с любыми желаемыми характеристиками. Делается это так. Например, мы хотим создать молекулу РНК, которая способна безошибочно узнавать вещество X и связываться с ним. Для этого синтезируют большое количество разных цепочек РНК, соединяя рибонуклеотиды друг с другом в случайном порядке. Раствор, содержащий полученную смесь молекул РНК, наливают на поверхность, покрытую веществом X. После этого остается лишь отобрать и исследовать те молекулы РНК, которые прилипли к поверхности. Технология незамысловата, но она действительно работает. Примерно таким способом получены рибозимы, катализирующие синтез нуклеотидов, присоединяющие аминокислоты к РНК и выполняющие множество других биохимических функций. Стирая грань между живым и неживым, уже растут на искусственных средах в лабораториях возмутительнейшие объекты – колонии размножающихся молекул РНК, способные к тому же синтезировать белки (правда, без этих самых белков – ферментов – заставить их расти пока не удается). Весомый вклад в эти исследования вносят ученые из Института химической биологии и фундаментальной медицины (г. Новосибирск) и Института белка (г. Пущино) под руководством академиков В. В. Власова и А. С. Спирина. Любопытно, что многие рибозимы работают лучше всего при низких температурах, иногда даже ниже точки замерзания воды – в крошечных полостях льда, где достигаются высокие концентрации реагентов. Некоторые считают это свидетельством того, что жизнь зарождалась при низких температурах.
Конечно, нельзя сказать, что в теории РНК-мира совсем нет проблем и трудностей. Их очень много. Основная, пожалуй, состоит в том, что большинство рибозимов очень малоэффективны по сравнению со своими белковыми аналогами. Во многих случаях это не принципиально, потому что на ранних этапах становления жизни белков еще не было, рибозимам не с кем было конкурировать, они на тот момент были "последним словом науки и техники". Их эффективности вполне хватало для того, чтобы предоставить химическим циклам, в которых они участвовали как катализаторы, решающее преимущество в скорости. Но некоторые ограничения могли оказаться принципиальными. В первую очередь это относится к вышеупомянутой способности рибозимов катализировать синтез собственных копий. В действительности тут все не так просто и гладко. В ныне живущих организмах таких рибозимов не обнаружено. Методом "искусственной эволюции" (то есть путем синтеза множества случайных последовательностей рибонуклеотидов, последующего отбора удачных вариантов, внесения в них небольших случайных изменений, нового отбора и т. д.) удалось получить рибозимы, которые худо-бедно могут "сшить" друг с другом два рибонуклеотида, но не всякие и не всегда. Осуществить полноценное копирование длинной молекулы РНК (то есть служить настоящими РНК-зависимыми РНК-полимеразами) они не могут. Чтобы изготовить из молекул РНК рибозим, который можно с полным правом назвать РНК-зависимой РНК-полимеразой, ученым пришлось воспользоваться наряду с "искусственной эволюцией" еще и сознательным, разумным планированием. В конце концов это удалось сделать – искомый рибозим был составлен из нескольких разных молекул РНК. Но даже и этот с таким трудом разработанный и изготовленный комплексный рибозим работает из рук вон плохо. В чем же дело?

Схема транспортной РНК. А, G, С, U – стандартные нуклеотиды аденозин, гуанозин, цитидин и уридин, другими буквами отмечены нестандартные (модифицированные) нуклеотиды, в том числе I – инозин. Как и многие другие молекулы РНК, тРНК образует петли за счет того, что рибонуклеотиды попарно "склеиваются друг с другом по принципу комплементарности. Как можно видеть на рисунке, С (цитозин) обычно склеивается с G (гуанином), а А (аденин) – с U (урацилом). Но в биологии, как мы уже знаем, нет правил без исключений: в двух местах этой молекулы, вопреки принципу комплементарности, G соединяется с U.
Не исключено, что дело тут в необходимых кофакторах – ионах металлов, в том числе редких. Известно, что очень многие белковые ферменты используют ионы металлов в качестве своих необходимых составных частей. Такие белки называют металлопротеинами. Похоже, это было справедливо и для рибозимов РНК-мира. До самого недавнего времени эту возможность упускали из виду. Однако недавно было обнаружено, что те самые искусственно полученные рибозимы, которые способны кое-как синтезировать РНК на РНКовой матрице, являются металлорибозимами: в их активном центре присутствует ион магния. Точно так же и проблему взаимодействия РНК с липидными мембранами удалось решить только благодаря объединению РНКовых комплексов с ионами металла, на этот раз – кальция.
Известно, что в древнем океане было гораздо больше, чем теперь, ионов различных тяжелых металлов, в том числе довольно экзотических, таких как вольфрам, молибден или ванадий[17]. Эти ионы до сих пор используются в качестве кофакторов многими белками, особенно у архаичных микроорганизмов. Может быть, дело у исследователей РНК-мира пойдет лучше, если они дадут рибозимам возможность "пользоваться услугами" ионов редких металлов?
У микроба ферроплазмы почти все белки содержат железо.
О важной и еще не до конца понятой роли металлов в жизни примитивных клеток свидетельствуют результаты изучения необычного микроба, обнаруженного в 2000 году в биореакторе металлургического завода в Туле.
Этот микроб, относящийся к надцарству архей (Archaea), получил название Ferroplasma acidiphilum. В отличие от большинства других архей и бактерий ферроплазма лишена жесткой клеточной стенки и размножается почкованием.
Ферроплазма живет в очень кислых водах, насыщенных растворенным железом и другими металлами. В природе такие условия встречаются в окрестностях месторождений сульфидных руд (например пирита). Ферроплазма сама производит органику из углекислого газа (подобно растениям), а питаться готовыми органическими веществами не может. В отличие от растений источником энергии для синтеза органики из СO2 ферроплазме служит не солнечный свет, а химическая реакция окисления двухвалентного железа (Fe2+=>Fe3+).

Ферроплазма в процессе почкования (почки показаны стрелками). По фотографии из статьи с первоописанием микроба (О. V. Golyshina et.al., 2000).
В ходе дальнейших исследований у ферроплазмы обнаружилось еще несколько необычных особенностей. В частности, оказалось, что один из ее ферментов (альфа-глюкозидаза) является железосодержащим металлопротеином, в то время как у всех остальных организмов ферменты этого класса никакого железа не содержат. Это побудило команду микробиологов – первооткрывателей ферроплазмы проверить и остальные белки странного микроорганизма на предмет содержания в них железа и других металлов.
Результат оказался более чем удивительным. Из 189 белков, которые ученым удалось выделить из ферроплазмы, лишь 26 не содержали железа. Остальные 163 (86 %) оказались железосодержащими металлопротеинами. Самое удивительное, что аналоги большинства из этих белков, встречающиеся у других организмов, не содержат железа.
Железо в белках ферроплазмы не является какой-то необязательной или второстепенной примесью. Исследователи показали, что аккуратное удаление железа из этих белков приводит к сильному изменению их структуры (денатурации) и потере функциональной активности. Похоже на то, что почти все белки ферроплазмы, по образному выражению авторов статьи, держатся на "железных заклепках".
Естественно, сразу возник вопрос о причинах такого обилия железосодержащих металлопротеинов у ферроплазмы. Возможно, это общее свойство всей группы микроорганизмов, к которой относится ферроплазма? Или, может быть, оно характерно именно для микробов, обитающих в кислых, насыщенных растворенным железом водах? Для ответа на этот вопрос исследователи выделили теми же методами металлосодержащие белки из ближайшего родственника ферроплазмы – архей Picrophilus torridus, а также из неродственного, но обитающего в таких же условиях микроорганизма – бактерии Acidithiobacillus ferrooxidans. Из этих микробов удалось выделить лишь 29 и 28 металлопротеинов соответственно, из которых только половина содержала железо. Самое главное, что все железосодержащие металлопротеины этих двух микробов оказались обычными, широко распространенными металлопротеинами, которые и у многих других организмов тоже содержат железо.
Тогда исследователи высказали дерзкую гипотезу. Они предположили, что ситуация, наблюдаемая у ферроплазмы, является случайно сохранившимся отголоском древнейших этапов развития жизни. Жизнь могла зародиться в микрополостях кристаллов пирита, в условиях, очень близких к тем, в которых ныне обитает ферроплазма. Главными отличиями этих микрополостей являются кислая среда и изобилие растворенного железа, которое в других, менее экзотических местообитаниях обычно в большом дефиците.
Древнейшие формы жизни поначалу активно использовали для осуществления необходимых химических реакций простые неорганические катализаторы, в первую очередь соединения железа и серы. Постепенно эти катализаторы замещались более эффективными органическими – то есть белками, и вполне естественно предположить, что первые белки включали в себя атомы железа как неотъемлемые структурные и функциональные компоненты. В дальнейшем, когда живые организмы стали осваивать другие местообитания, они сразу же столкнулись с резким дефицитом доступного железа, и отбор стал способствовать замене старых железосодержащих белков другими, не нуждающимися в железе для выполнения своих функций. В конце концов железо сохранилось только в тех белках, которые без него совсем уж никак обойтись не могут.
По мнению исследователей, обилие железосодержащих металлопротеинов у ферроплазмы объясняется тем, что предки этого микроба никогда не покидали кислых, богатых железом вод, и вся их эволюция протекала в условиях железного изобилия. Другие микроорганизмы, обитающие сегодня в похожих условиях, вероятно, попали туда вторично и на каких-то этапах своей эволюции сталкивались с дефицитом железа. Те немногочисленные белки ферроплазмы, в которых железа все-таки нет, возможно, достались ей от этих новых соседей в результате горизонтального (межвидового) обмена генами, что у микробов – обычное дело (см. главу "Наследуются ли приобретенные признаки").
Впрочем, есть одно обстоятельство, заставляющее усомниться в столь глубокой древности ферроплазмы и ее металлопротеинов. Дело в том, что этот микроорганизм является аэробным, для окисления железа ему необходим кислород, тогда как древнейшие этапы эволюции жизни протекали в бескислородных условиях (см. главу "Планета микробов").
Как РНК решала энергетический вопрос. Чтобы два рибонуклеотида соединились вместе, образовав маленькую молекулу РНК, к одному из них должен быть присоединен дополнительный фосфат (или сразу два). Получившаяся молекула – рибонуклеотид с лишним фосфатом – обладает большим количеством энергии, которая при наличии подходящих катализаторов может быть использована для выполнения разных полезных "работ". В том числе для соединения двух рибонуклеотидов в одну молекулу – маленькую РНК. Рибонуклеотиды с дополнительными фосфатами первоначально использовались, скорее всего, только как "строительные кирпичики" при синтезе РНК. Кирпичики, надо сказать, очень удобные – ведь они включают в себя не только строительный материал, но еще и энергию, необходимую для выполнения строительных работ! Впоследствии они стали использоваться для тысяч других важных дел – везде, где для выполнения какой-то работы требуется энергия. Все живое и по сей день пользуется фосфорилированными рибонуклеотидами как универсальными поставщиками энергии при выполнении энергоемких задач. Самая известная из этих "энергетических" молекул – АТФ (аденозинтрифосфат). Это обычный рибонуклеотид, к которому присоединены два дополнительных фосфата. АТФ одновременно и источник энергии для множества энергоемких реакций, и один из кирпичиков для синтеза РНК. Так земная жизнь нашла универсальное решение сразу двух задач: запасания энергии в удобной форме и синтеза РНК – главных молекул жизни.

НАД (никотинамид-аденин-динуклеотид) – важнейший участник энергетического метаболизма клетки, используемый как универсальный переносчик протонов и электронов. Представляет собой два соединенных рибонуклеотида: обычный (аденозин) и "нестандартный" (никотинамид). Структура этого вещества – еще одно "напоминание* об эпохе РНК-мира.
Оболочка
Все организмы дискретны в пространстве и имеют наружную оболочку. Трудно представить себе живое существо в виде туманного облачка или раствора (разве что в фантастическом романе). Однако поначалу преджизнь существовала именно в виде растворов. Чтобы не раствориться окончательно, не рассеяться в водах древних водоемов, "живые растворы" должны были ютиться в крошечных полостях, которые часто встречаются в минералах. Это тем более удобно, что некоторые минералы (например, пирит) являются неплохими катализаторами для многих биохимических реакций. Кроме того, поверхность минералов могла служить своеобразной матрицей, основой, к которой прикреплялись молекулы РНК. Упорядоченная структура кристаллов помогала упорядочить и структуру этих молекул, придать им нужную пространственную конфигурацию.
Но рано или поздно преджизнь должна была обзавестись собственными оболочками – перейти от доорганизменного уровня к организменному. Идеальным материалом для таких оболочек являются липиды (жиры), молекулы которых способны образовывать на поверхности воды тончайшие пленки. Если взболтать такую воду, в ее толще образуется множество мелких пузырьков – водяных капелек, покрытых двухслойной липидной оболочкой (мембраной). Эти капельки проявляют интересные свойства, которые делают их похожими на живые клетки. Например, они способны осуществлять обмен веществ. Липидные мембраны обладают избирательной проницаемостью: одни молекулы сквозь них проходят, другие – нет. Благодаря этому одни вещества втягиваются в каплю, другие выводятся, третьи – накапливаются внутри. Правда, для того, чтобы это происходило постоянно, одних мембран недостаточно. Нужно еще, чтобы внутри капли одни вещества превращались в другие, а для этого там должны находиться катализаторы – белки или РНК.
Изучением свойств водно-липидных капель (коацерватов) занимался академик А. И. Опарин. Он считал, что коацерваты были одним из этапов на пути возникновения жизни. Опарин обнаружил, что при определенных условиях коацерваты могут расти и даже "размножаться" делением[18].
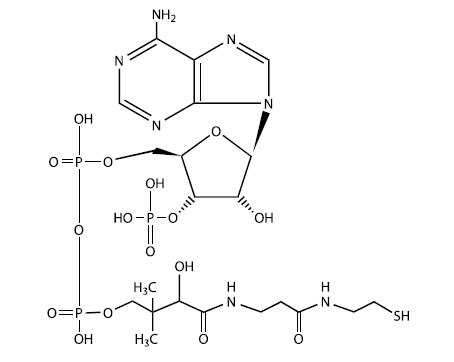
Кофермент А – одна из важнейших биоактивных молекул живой клетки – представляет собой модифицированный рибонуклеотид аденозин. К одному из остатков фосфорной кислоты присоединен "хвост", отдаленно напоминающий очень короткую белковую молекулу. Хвост заканчивается группой – SH, которая является активным центром молекулы. К этой сере может присоединяться ацетильная группа (-СО-СН3)у которая затем будет использована, например, для синтеза жирных кислот (ключевой этап синтеза липидов). Строение молекулы кофермента А заставляет задуматься о древнейших этапах эволюции жизни, когда рибонуклеотиды и простейшие белки (пептиды) еще только учились «жить и работать вместе»
Первые коацерваты могли образоваться самопроизвольно из липидов, синтезированных абиогенным путем. Впоследствии они могли вступить в симбиоз (взаимовыгодное сожительство) с "живыми растворами" – колониями самовоспроизводящихся молекул РНК, среди которых были и рибозимы, катализирующие синтез липидов. Подобное сообщество уже можно назвать организмом. У всех живых существ до сих пор в синтезе липидов важнейшую роль играет кофермент А, представляющий собой не что иное, как модифицированный рибонуклеотид. Это еще одно напоминание об РНК-мире.
Камнем преткновения для теории РНК-мира в течении некоторого времени была неспособность молекул РНК эффективно взаимодействовать с липидными мембранами. Недавно, однако, было показано, что комплексы из нескольких разных молекул РНК и ионов кальция способны не только прикрепляться к мембранам, но и регулировать их проницаемость.
РНК обзаводится помощниками
В дальнейшем РНК-организмы приобрели несколько важных усовершенствований. Самое главное из них состояло в том, что они научились синтезировать аминокислотные полимеры – сначала короткие пептиды, а затем и длинные белки. Эти вещества стали для РНК-организмов универсальными помощниками, справляющимися с большинством биологических "работ" гораздо лучше, чем рибозимы.
Откуда же взялась у РНК-организмов способность синтезировать белки? Чтобы ответить на этот вопрос, мы должны поближе познакомиться с рибосомами – сложными молекулярными "машинками", при помощи которых синтезируют белки все современные живые клетки.
Рибосомы у всех живых существ – от бактерий до человека – устроены очень похоже. По-видимому, это означает, что рибосомы в их "современном" виде имелись уже у общего предка всех нынешних форм жизни – у Луки, о котором говорилось в начале главы. Рибосома состоит из двух частей, или субъединиц, – большой (главной) и малой (вспомогательной). Основу обеих субъединиц составляют молекулы рибосомной РНК (рРНК). Снаружи к молекулам рРНК прилегают молекулы рибосомных белков. Поскольку рибосомы играют главную роль в синтезе белка (трансляции), вопрос о происхождении синтеза белка фактически сводится к вопросу о происхождении рибосом.
До самого недавнего времени многим экспертам казалось, что загадка происхождения рибосом вряд ли когда-нибудь будет разгадана. Ведь в природе не осталось никаких "переходных звеньев", то есть более простых молекулярных комплексов, которые могли бы претендовать на роль "предков" рибосом. Однако в начале 2009 года канадские биохимики, похоже, нашли ключик к этой тайне в самой структуре рибосом современных организмов[19].
Они сосредоточились на самой главной части рибосомы – на молекуле РНК, которая называется 23S-pPHK и является основой большой субъединицы рибосомы. Эта молекула весьма велика: она состоит почти из 3000 нуклеотидов. В клетке она сворачивается в сложный трехмерный "клубок". Разные петли, выступы и другие элементы структуры этого "клубка" обеспечивают выполнение разных функций: связь с рибосомными белками, присоединение малой субъединицы, присоединение и удерживание в нужных позициях молекул транспортных РНК (тРНК), которые несут на своих "хвостиках" аминокислоты, необходимые для синтеза белка.
Проведенные ранее эксперименты показали, что рибосомные белки играют в рибосоме вспомогательную роль: они делают ее более стабильной и повышают эффективность ее работы, однако все главные действия, необходимые для синтеза белка, осуществляются не белками, а рибосомными РНК. Это значит, что изначально рибосомы могли состоять только из рРНК, а белки добавились позже. Самый главный этап трансляции – присоединение аминокислот к синтезируемой белковой молекуле – осуществляется молекулой 23S-pPHK. Поэтому логично предположить, что все началось именно с этой молекулы.
Однако молекула 23S-pPHK слишком велика и сложна, чтобы появиться в готовом виде в результате случайного комбинирования нуклеотидов. Таким образом, ключевой допрос состоит в том, могла ли 23S-pPHK произойти от более простой молекулы-предшественницы в результате постепенной эволюции, то есть путем последовательного добавления новых фрагментов. Ученым удалось показать, что структура 23S-pPHK свидетельствует именно о таком ее происхождении.
Целостность трехмерной структуры молекулы 23S-pPHK поддерживается разнообразными связями между ее участками. Некоторые части молекулы сворачиваются в двойные спирали. К двойным спиралям "приклеиваются" другие участки молекулы, состоящие из нескольких идущих подряд аденозинов. Связи, возникающие между двойными спиралями и "стопками" аденозинов, необходимы для поддержания стабильной трехмерной структуры той части молекулы, к которой принадлежит аденозиновая "стопка", но они не влияют на стабильность той ее части, к которой принадлежит двойная спираль. Иными словами, если мы разорвем какую-нибудь из этих связей, это нарушит структуру той части молекулы, где находится аденозиновая "стопка", но не причинит вреда той части, где расположена двойная спираль. Таким образом, если 23S-pPHK развивалась постепенно из простой молекулы-предшественницы, то сначала должны были появляться двойные спирали, и только потом к ним могли "пристраиваться" аденозиновые стопки.
Изучая структуру 23S-pPHK, исследователи обратили внимание, что в одной части молекулы имеется скопление двойных спиралей и почти нет аденозиновых стопок. Это наблюдение навело ученых на мысль, что эволюция молекулы 23S-pPHK могла начаться именно с этого фрагмента молекулы.
Но если этот фрагмент был той "затравкой", с которой началась эволюция 23S-pPHK, то следует ожидать, что именно в нем находится какой-то важный функциональный центр молекулы. Так ли это? Оказывается, это действительно так: именно этот участок молекулы 23S-pPHK играет ключевую роль в присоединении аминокислот к синтезируемому белку. Он удерживает в правильных позициях "хвосты" двух молекул тРНК (той, что принесла предыдущую аминокислоту, уже присоединенную к белку, и той, что принесла следующую аминокислоту). Именно этот участок молекулы обеспечивает сближение новой аминокислоты с предыдущей, уже присоединенной к белку, и катализирует соединение аминокислоты с белком.
Обнаружив эти факты, исследователи перешли к более тонкому анализу структуры 23S-pPHK. Они подразделили молекулу на более тонкому анализу. Они подразделили молекулу на 60 относительно самостоятельных структурных блоков и детально проанализировали характер связей между ними. Фактически они рассматривали молекулу как сложный трехмерный "пазл" и пытались выяснить, поддается ли он сборке и разборке без поломки деталей. Оказалось, что молекулу действительно можно постепенно "разобрать", ни разу не нарушив структуру остающихся блоков. Сначала можно отделить 19 блоков, причем структура оставшихся блоков остается неповрежденной. После этого отделяются еще 11 блоков, затем еще 9, 5, 3, 3, 2, 2, 2; наконец, еще три блока можно отделить последовательно по одному. После этого остается "неразобранным" лишь маленький фрагмент молекулы, составляющий 7 % от ее общей массы. Этот неразобранный фрагмент представляет собой тот самый каталитический центр, ответственный за удерживание двух молекул тРНК и присоединение аминокислот к белку.
Возможность последовательной разборки молекулы без повреждения остающихся частей – факт весьма нетривиальный. Все блоки молекулы связаны друг с другом, причем связи эти имеют направленный характер: при их разрыве один блок повреждается, а другой – нет. Можно представить систему блоков и связей между ними как множество точек, соединенных стрелками, причем стрелка будет указывать на тот блок, который повреждается при разрыве связи. Если бы эти стрелки образовали хотя бы одну кольцевую структуру (иными словами, если бы мы, двигаясь из какой-то точки по стрелкам, могли вернуться в ту же точку), то разобрать молекулу без повреждения остающихся частей было бы невозможно. Однако ни одной такой кольцевой структуры в молекуле 23S-pPHK не обнаружилось. Если бы направление связей было случайным, вероятность отсутствия кольцевых структур составляла бы менее одной миллиардной. Значит, это вряд ли результат случайности. По-видимому, структура связей между блоками молекулы отражает последовательность добавления этих блоков в ходе постепенной эволюции молекулы.
Получается, что исходной функциональной молекулой – "проторибосомой", с которой началась эволюция рибосомы, – был каталитический центр молекулы 23S-pPHK, ответственный за соединение аминокислот.
Могла ли такая "проторибосома", способная удерживать две молекулы тРНК и сближать в пространстве прикрепленные к ним аминокислоты, выполнять какую-то полезную функцию в РНК-организме? Эксперименты позволяют ответить на этот вопрос утвердительно. Методом искусственной эволюции были получены функциональные РНК (рибозимы), способные катализировать соединение аминокислот, прикрепленных к тРНК, в короткие белковые молекулы. Структура этих искусственно выведенных рибозимов очень близка к структуре той проторибосомы, которую "вычислили" канадские биохимики на основе изучения структуры 23S-pPHK.
По-видимому, проторибосома была просто устроенным рибозимом, катализирующим синтез небольших белковых молекул в РНК-организме. Специфичность синтеза поначалу была очень низкой (аминокислоты выбирались более или менее случайно). В дальнейшем к проторибосоме добавлялись новые блоки, причем добавлялись они таким образом, чтобы не нарушить структуру активного центра молекулы, а также всех тех блоков, которые присоединились ранее. Если очередная мутация приводила к нарушению уже сложившихся структур, она отсеивалась отбором.
Ученые детально реконструировали предполагаемый процесс постепенной эволюции 23S-pPHK. Первые восемь дополнительных блоков присоединились к проторибосоме таким образом, что образовали нечто вроде массивного "основания", благодаря которому структура проторибосомы стала гораздо более стабильной. Следующие 12 блоков еще более укрепили и расширили это "основание". Новые блоки образовали поверхность контакта с малой субъединицей, что позволило включить ее в состав рибосомы. В числе последних добавились блоки, образующие особые выросты на поверхности большой субъединицы. Функция этих выростов состоит в том, что они помогают рибосоме выбирать "правильную" тРНК, несущую нужную аминокислоту, а также выпускать из рибосомы "отработанные" тРНК. В итоге проторибосома оказалась окружена другими блоками со всех сторон за исключением канала, который был оставлен для выхода образующейся белковой молекулы.
Таким образом, 23S-pPHK, при всей ее кажущейся сложности, построена на основе довольно простого принципа. Ее блочная структура свидетельствует о том, что она могла довольно быстро развиться в ходе эволюции из проторибосомы под действием мутаций и отбора.
ДНК и РНК – хранители наследственной информации. ДНК, как и РНК, формируются из кирпичиков – нуклеотидов, только чуть-чуть других. Чтобы из РНК-кирпичика (рибонуклеотида) сделать ДНК-кирпичик (дезоксирибонуклеотид), достаточно одной простой реакции – отнять у рибозы один из атомов кислорода. Это придает молекуле стабильность, а заодно лишает ее способности совершать активные действия.
Молекулы ДНК (как и РНК) способны к самокопированию, правда, для этого нужны катализаторы – белки или рибозимы. Наследственная информация, хранящаяся в ДНК в виде последовательности нуклеотидов, может "переписываться" на РНК (так создаются матричные РНК, мРНК) и обратно. Точность копирования обеспечивается в значительной мере автоматически – благодаря особому свойству нуклеотидов, которое называют свойством комплементарности: против каждого нуклеотида исходной молекулы (матрицы) в синтезируемой копии (реплике) может встать только один, строго определенный нуклеотид из четырех возможных. Напротив гуанина (Г всегда становится цитозин (Ц), напротив урацила (У) или замещающего его в молекуле ДНК тимина (Т) – только аденин (А). Когда на этой реплике синтезируется новая реплика, она окажется точной копией исходной молекулы.
Фрагмент двойной цепи ДНК. По краям – "скелеты" одиночных цепей, составленные из остатков дезоксирибозы и фосфорной кислоты. В центре – две пары азотистых оснований, соединенных друг с другом по принципу комплементарности. Между аденином (А) и тимином (Т) образуются две водородные связи, между гуанином (Г) и цитозином (Ц) – три. Поэтому аденин может склеиться только с тимином, а гуанин – только с цитозином.
"Взаимовыгодное сотрудничество" РНК и белков (пептидов), вероятно, складывалось постепенно. Можно предположить, что изначально химические циклы с участием РНК и пептидов формировались порознь, возможно, в несколько разных условиях. Об этом свидетельствует химический состав этих веществ. В РНК много фосфора: остатки фосфорной кислоты вместе с остатками рибозы составляют "скелет" молекулы. Сера в состав РНК не входит. В белках, наоборот, нет фосфора, зато имеется сера, которая играет весьма важную роль в поддержании пространственной структуры белковой молекулы.
Поначалу синтез белков, осуществлявшийся РНК-организмами, скорее всего, не был строго специфичным: последовательности аминокислот из раза в раз воспроизводились не точно, а лишь приблизительно. Поскольку точность в данном случае резко повышала жизнеспособность организмов, естественный отбор способствовал выработке все более точных методов синтеза белка. Дело кончилось возникновением универсальной системы очень точного синтеза любого требуемого пептида. Это и был генетический код вкупе с рибосомами "современного" типа.
Генетический код – универсальный для всех живых существ способ, посредством которого первичная структура белковой молекулы (последовательность аминокислот) "кодируется" в молекуле ДНК (или РНК). Каждая аминокислота кодируется тремя нуклеотидами (кодоном, или триплетом). Нуклеотидов в ДНК всего 4, поэтому они могут образовывать 64 разных триплета. Аминокислот в белках всего 20, поэтому генетический код "избыточен": многие аминокислоты кодируются не одним, а несколькими взаимозаменимыми кодонами. Считывание генетической информации происходит в два этапа. Сначала информация "переписывается" с ДНК на РНК (транскрипция). Эту операцию осуществляет специальный фермент – ДНК-зависимая РНК-полимераза. Полученная в результате транскрипции молекула РНК, содержащая "инструкцию" по синтезу белка, называется матричной РНК (мРНК). Выполнение этой "инструкции", то есть синтез белка (трансляция), осуществляется рибосомами.
Вторым крупным усовершенствованием РНК-организмов было приобретение ДНК. Молекулы ДНК более устойчивы, чем РНК, и потому являются более надежными хранителями наследственной информации. Платой за стабильность стала неспособность молекул ДНК сворачиваться в сложные трехмерные структуры и выполнять какие-либо активные действия. Изначально ДНК, скорее всего, была чем-то вроде покоящейся фазы в жизненном цикле самовоспроизводящихся колоний РНК, и лишь много позднее она стала основным носителем наследственной информации.
Формы существования наследственной информации. Наследственная (генетическая) информация может существовать в двух формах – в виде ДНК и РНК. Копированием и переписыванием этой информации занимаются особые ферменты – НК-полимеразы. Существует четыре типа НК-полимераз:
1. ДНК-зависимые ДНК-полимеразы – осуществляют репликацию ДНК, то есть синтезируют ДНК на матрице ДНК. Эти ферменты просто копируют молекулы ДНК, как на ксероксе.
2. ДНК-зависимые РНК-полимеразы – осуществляют транскрипцию, то есть синтезируют РНК на матрице ДНК.
3. РНК-зависимые РНК-полимеразы – осуществляют репликацию РНК, то есть синтезируют РНК на матрице РНК.
4. РНК-зависимые ДНК-полимеразы (обратные транскриптазы, ревертазы) – осуществляют обратную транскрипцию, то есть синтезируют ДНК на матрице РНК.
Есть веские основания полагать, что первыми появились ферменты третьего типа, а от них потом произошли все остальные типы НК-полимераз.
Наследие РНК-мира
В последние годы одним из самых быстро развивающихся направлений в молекулярной биологии стало исследование разнообразных маленьких молекул РНК, которые, как выяснилось, играют огромную роль в жизни клетки. В результате этих исследований представления о молекулярных основах жизни сильно изменились. Еще лет 10–15 назад казалось, что РНК играет в клетке все-таки второстепенную роль. Сегодня стало ясно, что молекулы РНК являются активными участниками множества жизненно важных процессов. Постоянно открываются новые функциональные молекулы РНК и новые "роли", выполняемые этими молекулами в клетке. Эти открытия очень хорошо согласуются с теорией РНК-мира. Действительно, если древнейшие живые организмы умели обходиться вообще без белков и ДНК и все функции в них выполнялись молекулами РНК, то можно ожидать, что и в современных организмах эти многофункциональные молекулы не остались без работы (см. также главу "На подступах к неведомому").
Одним из явных отголосков эпохи РНК-мира являются недавно открытые удивительные структуры, получившие название РНК-переключателей.
РНК-переключатели впервые были обнаружены в 2002 году Рональдом Брейкером и его коллегами из Йельского университета. С тех пор число публикаций, посвященных этому странному и очень древнему механизму генной регуляции, стремительно растет.
Работа гена начинается с транскрипции – создания молекулы мРНК на матрице ДНК. Транскрибируется не только та часть ДНК, которая кодирует белок, но и кое-что "лишнее", в том числе участок перед началом кодирующей области. Здесь-то и располагаются РНК-переключатели. Они представляют собой последовательности нуклеотидов, которые сразу после транскрипции сворачиваются в замысловатые трехмерные структуры. Сворачивание осуществляется на основе принципа комплементарности (так же, как это происходит, например, с транспортными и рибосомными РНК). Самое важное, что область, где находятся РНК-переключатели, транскрибируется первой. РНК-переключатели приходят в рабочее состояние – то есть принимают нужную конфигурацию – сразу, как только их транскрибировали, и задолго до того, как закончится транскрипция всего гена. Это позволяет им прервать транскрипцию и тем самым фактически выключить ген.
РНК-переключатель состоит из двух функциональных частей. Первая часть представляет собой весьма избирательный и чувствительный рецептор, который способен связываться с одной строго определенной молекулой (например, с аминокислотой глицином или с S-аденозилметионином). Вторая часть устройства – это собственно переключатель. Когда рецептор связывается со "своей" молекулой, переключатель меняет свою пространственную конфигурацию, что и приводит к изменению активности гена. Например, переключатель может образовать "шпильку" – торчащий двухнитевой участок, который блокирует дальнейшую транскрипцию и на котором недоделанная информационная РНК просто-напросто обрывается.
Ключевой молекулой, которая приводит в действие РНК-переключатель, часто является вещество, производимое белком, ген которого этим переключателем регулируется. Например, если продуктом гена является белок, синтезирующий вещество А, то РНК-переключатель этого гена с большой вероятностью будет реагировать именно на вещество А. Таким образом формируется отрицательная обратная связь: когда какого-то продукта становится слишком много, производство белка, синтезирующего этот продукт, приостанавливается.
РНК-переключатели широко распространены во всех трех надцарствах живой природы – у бактерий, архей и эукариот. Наиболее разнообразны они у бактерий. Поскольку открыты они были всего несколько лет назад, неудивительно, что почти каждый месяц мы узнаем о них что-то новое. Сначала думали, что все РНК-переключатели снижают активность генов, но вскоре среди них были открыты и активаторы. Думали, что регуляторные контуры с участием РНК-переключателей всегда просты: один ген – один переключатель – одно сигнальное вещество. Однако в 2006 году в журнале Science появилась статья группы американских исследователей во главе с Брейкером, в которой описан новый тип регуляторного РНК-устройства, состоящего из двух разных РНК-переключателей[20]. Ученые установили, что комплекс из двух переключателей работает как логический элемент NOR (ИЛИ-НЕ). Иными словами, ген выключается, если оба или хотя бы один из двух переключателей свяжется со своей молекулой.
Открытие показало, что возможности безбелковой РНК-регуляции активности генов далеко не так ограниченны, как думали раньше. На основе простых РНК-переключателей могут создаваться более сложные регуляторные устройства, способные учитывать сразу несколько параметров окружающей среды.
Уже первооткрывателям РНК-переключателей сразу стало ясно, что они столкнулись с чем-то чрезвычайно древним.
Человек, обладающий хорошим воображением, может представить себе эту картину в красках – "считываемый" ген вдруг начинает шевелиться, воспринимать сигналы из окружающей среды, реагировать на них и вмешиваться в работу считывающего устройства: не читай меня больше! Таким образом, становится понятно, как далеки от истины были исходные представления об РНК как о безынициативном посреднике между ДНК и машиной синтеза белка.
В начале было сообщество?
Многие биологи полагают, что все разнообразие жизни на нашей планете происходит от единственного исходного вида – "универсального предка" по имени Лука (об этом прародителе всего живого на Земле мы уже упомянули в начале главы). Другие, в том числе крупнейший микробиолог академик Г.А. Заварзин, с этим не согласны. Они исходят из того, что устойчивое существование биосферы возможно только при условии относительной замкнутости биогеохимических циклов – в противном случае живые существа очень быстро израсходуют все ресурсы или отравят себя продуктами собственной жизнедеятельности.
Замкнутость циклов может быть обеспечена только сообществом из нескольких разных видов микроорганизмов, разделивших между собой биогеохимические функции. Одни, используя ресурсы среды, наполняют ее продуктами своей жизнедеятельности, а другие, используя эти продукты, возвращают в качестве своих отходов первоначальный ресурс во внешнюю среду. Примером такого сообщества являются циано-бактериальные маты, о которых пойдет речь в главе 3. Г.А. Заварзин считает, что организм, способный в одиночку замкнуть круговорот, так же невозможен, как и вечный двигатель.
Для этапа химической преджизни это еще более очевидно. Никакая отдельно взятая органическая молекула не сможет устойчиво самовоспроизводиться и поддерживать гомеостаз в окружающей среде. На это способны только комплексы из довольно большого числа разных молекул, между которыми наладилось "взаимовыгодное сотрудничество" (симбиоз).
Если принять этот ход рассуждений, то окажется, что, скорее всего, Лука был не единым видом микроорганизмов, а полиморфным сообществом, в котором происходил активный обмен наследственным материалом между организмами. Разнообразие, симбиоз, разделение функций и информационный обмен – изначальные свойства земной жизни.
Впрочем, следует помнить, что в биологии не бывает правил без исключений и даже видимая безупречность логических рассуждений не всегда может служить критерием истины. Теоретически все-таки можно себе представить единичный вид микроорганизмов, существующий на планете в течение очень долгого времени. Например, если "пищей" ему служат какие-либо вещества, поступающие понемножку из земных недр, а отходы жизнедеятельности либо перерабатываются в геохимических круговоротах без участия живых организмов, либо, к примеру, просто захораниваются в земной коре. Таким образом, этот гипотетический микроорганизм попросту встраивается в уже существующий геохимический цикл, лишь ускоряя его.
Однако в целом на сегодняшний день утверждение "в начале было сообщество" представляется, на мой взгляд, более вероятным, чем "в начале был один вид микробов".
В следующей главе мы подробнее поговорим о микробах, микробных сообществах и о самом долгом из всех этапов истории Земли, в течение которого микробы были господствующей формой жизни.
Что почитать на эту тему в Интернете
В. В. ВЛАСОВ, А. В. ВЛАСОВ. Жизнь начиналась с РНК // Наука из первых рук. № 2(3). 2004. С. 6–19. http://evolbiol.ru/vlasov.htm
Г. А. ЗАВАРЗИН. Становление системы биогеохимических циклов. // Палеонтологический журнал. № 6. 2003. С. 16–24. http://evolbiol. ru/zavarzin2003.htm
А. В. МАРКОВ. Обзор "Зарождение жизни. Прокариотная биосфера" 2003–2007. http://evolbiol.ru/paleobac.htm
A. Ю. РОЗАНОВ. Цианобактерии и, возможно, низшие грибы в метеоритах. 1996. http://www.pereplet.ru/obrazovanie/stsoros/203.html
B. Н. СНЫТНИКОВ, В. Н. ПАРМОН. Жизнь создает планеты? // Наука из первых рук. № о. 2004. С. 20–31. http://evolbiol.ru/npr_snytnikov.pdf
М. А. ФЕДОНКИН. Сужение геохимического базиса жизни и эвкариотнзация биосферы: причинная связь // Палеонтологический журнал. № 6. 2003. С. 33–40.
М. А. ФЕДОНКИН. Геохимический голод и становление царств // Химия и жизнь, http://elementy.ru/lib/25583/25585