Глава 1. Генетическая информация
Ключевой проблемой биологии, по-видимому, можно считать вопрос о том, как увековечивает свой опыт живая материя.
При изучении любого биологического феномена необходимо знание основных генетических закономерностей. Особенно это относится к изучению поведения. Спор о роли наследственности прошел через историю всех «поведенческих» наук. Во многом причиной этому являлось непонимание спорящими сторонами законов генетики – методологической основы современной биологии.
1.1. Понятие генетической информации
Как хранится и воспроизводится генетическая информация? Какие структуры организма являются ее «хранилищем»? Как генетическая информация проявляется и как изменяется в ряду поколений? Без понимания этих вопросов немыслимо подходить к изучению любого аспекта поведения.
Носителями наследственной информации в природе являются нуклеиновые кислоты. Известно два вида нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). Генетическая информация записана на молекуле ДНК, которая служит местом ее хранения. Эта информация записана при помощи особых «букв» – нуклеотидов, которые являются мономерами ДНК.
Нуклеотиды ДНК представляют собой сложные молекулярные комплексы, состоящие из азотистого основания, углевода пентозы (дезоксирибоза) и остатка фосфорной кислоты. Четыре типа азотистых оснований (аденин – А; тимин – Т; гуанин – Г; цитозин – Ц) определяют специфичность четырех типов нуклеотидов ДНК. Таким образом, генетическая информация всех живых существ на Земле записана с помощью всего четырех «букв». Как такое возможно? Дело в том, что «слова», записанные этими «буквами», очень большие, что позволяет посредством всего четырех «букв» компоновать бесчисленное количество комбинаций. «Слова» генетического «словаря», которыми «описана» жизнь на Земле, называются генами.
Гены – единицы генетической информации. Иногда они небольшого размера, иногда состоят из тысяч нуклеотидов. Дать краткое и точное определение гену весьма сложно – это одна из центральных проблем генетики. Весь XX в. представления о гене непрерывно изменялись. В рамках классической генетики это было, скорее, абстрактное понятие элементарной структуры, отвечающей за определенный признак организма. После рождения молекулярной биологии под геном обычно понимался участок ДНК, кодирующий отдельный признак. Но последующие открытия «перекрывающихся» генов, «мозаичной» структуры, псевдогенов, мобильных генетических элементов, регуляторных последовательностей и многого другого все более усложняли и размывали понятие гена (Курчанов Н. А., 2009). С большой степенью условности остановимся на лаконичном определении гена, которое приводит в своей книге В. З. Тарантул: «Ген – это физическая (определенный участок ДНК) и функциональная (кодирует белок или РНК) единица наследственности» (Тарантул В. З., 2003).
Воспроизводится генетическая информация во время репликации. Репликацией называется процесс удвоения молекул ДНК. Именно уникальная способность ДНК к самоудвоению определяет ее ключевую роль в живом организме как носителя наследственности. Каждая молекула ДНК состоит из двух цепей нуклеотидов. Эти цепи соединены друг с другом по принципу комплементарности (А – Т, Г – Ц) при помощи водородных связей. Таким образом, информации цепей взаимозависимы: каждая цепь несет информацию о другой.
Начинается репликация с локального участка молекулы ДНК, где двойная спираль ДНК раскручивается, разрываются водородные связи между нуклеотидами соседних цепей и цепи расходятся. К образовавшимся свободным связям каждой цепи присоединяются по принципу комплементарности (А – Т, Г – Ц) свободные нуклеотиды, находящиеся в клетке. Этот процесс идет вдоль всей молекулы ДНК с участием сложного ферментного комплекса. Поскольку у каждой дочерней молекулы ДНК одна нить происходит от материнской молекулы, а другая является вновь синтезированной, данная модель репликации получила название «полуконсервативной» (рис. 1.1). Мы рассмотрели только сам принцип репликации, не вникая в исключительно сложные биохимические процессы.
Две новые молекулы ДНК представляют собой копии исходной молекулы. Полуконсервативный механизм репликации ДНК столь же универсален в природе для воспроизведения генетического материала, как и сама структура ДНК.
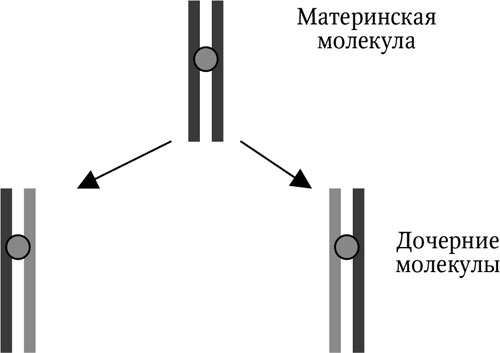
Рис. 1.1. Полуконсервативный принцип репликации ДНК
Другая нуклеиновая кислота – РНК, участвует в процессах реализации генетической информации. Ее нуклеотиды отличаются от нуклеотидов ДНК одним азотистым основанием (урацил вместо тимина) и пентозой (рибоза). В настоящее время считается, что РНК была первой «молекулой жизни», объединившей в себе множество функций. Только потом функция хранения генетической информации перешла к ДНК, а функция катализа – к белкам. Однако разнообразные виды РНК в той или иной мере сохраняют свои первоначальные способности.
Любая информация в организме должна не просто храниться, но и работать. Реализация генетической информации происходит в ходе экспрессии генов – в процессах транскрипции и трансляции.
Транскрипцией называется процесс переноса генетической информации с ДНК на РНК. Информация переписывается с языка ДНК-нуклеотидов на язык РНК-нуклеотидов (рис. 1.2).

Рис. 1.2. Комплементарность смысловой цепи ДНК и нити РНК в процессе транскрипции: А – аденин; Т – тимин; Г – гуанин; Ц – цитозин; У – урацил
Синтез РНК чаще происходит только на одной из двух нитей ДНК (смысловой) и не по всей ее длине, а лишь в пределах одного гена. Таким образом, ген можно представить как участок ДНК, несущий информацию об одной рибонуклеотидной цепи, но это определение осложняется множеством исключений. Так, данное определение не подходит к малым РНК.
В результате процесса транскрипции синтезируются все молекулы РНК (и-РНК, т-РНК, р-РНК и др.). Эти молекулы после биохимических преобразований направляются в цитоплазму для участия в другом важнейшем процессе – трансляции, который протекает на рибосомах.
Трансляцией называется процесс синтеза полипептидной цепочки на нити и-РНК. На и-РНК переписана генетическая информация о структуре белков, которая закодирована последовательностью нуклеотидов. Полипептидная нить из аминокислот, приносимых т-РНК, синтезируется на и-РНК. Этот процесс происходит на рибосомах. Сами рибосомы состоят из многочисленных белков и р-РНК. При трансляции информация переписывается с языка РНК-нуклеотидов на язык аминокислот. Этот процесс, один из самых сложных в природе, происходит в несколько этапов с участием многих ферментов.
Соответствие структуры гена (в нуклеотидах) и структуры кодируемого им белка (в аминокислотах) было названо колинеарностью. Колинеарность определяется генетическим кодом. Кодировка соответствия двух типов записи информации выражается в том, что каждая аминокислота в структуре белка зашифрована последовательностью из трех нуклеотидов – триплетом или кодоном (рис. 1.3). Расшифровка генетического кода, завершенная к 1965 г., явилась одной из триумфальных побед генетики.

Рис. 1.3. Колинеарность и-РНК и полипептида, синтезированного в процессе трансляции (в полипептиде показаны комплементарные триплетам аминокислоты)
Процессы репликации, транскрипции и трансляции получили в биологии название матричных, поскольку в них одна молекула служит матрицей (образцом) для синтеза другой. Матричные процессы являются определяющими для протекания жизни на Земле.
Порядок переписывания генетической информации в клетке определяет поток информации в живой природе:
ДНК → РНК → белок.
Такая направленность была названа одним из основателей молекулярной биологии Ф. Криком (1916–2004) центральной догмой молекулярной биологии. Хотя современная генетика знает и альтернативные пути, этот поток информации реализуется в подавляющем большинстве живых систем.
Единство генетического кода и общность потока генетической информации в живой природе свидетельствуют о единстве происхождения всех живых организмов на нашей планете.
1.2. Организация генетического материала
Структурно-функциональная организация генетического аппарата определяет деление всех живых организмов на прокариот и эукариот. У прокариот (к которым относятся бактерии и археи) ДНК представлена кольцевой молекулой и находится в цитоплазме клетки. У эукариот (к которым относятся все остальные организмы) структурными носителями генетической информации ДНК являются хромосомы, находящиеся в ядре.
Хромосомы представляют собой сложную многоуровневую структуру, в которой ДНК взаимодействует с различными белками. Базовым уровнем этой структуры являются нуклеосомы, представляющие собой глобулы из восьми молекул белков гистонов, обвитых ДНК. Нуклеогистоновая нить в дальнейшем многократно укладывается, формируя компактные хромосомы. Такая структура открывает широкие возможности для регуляции.
Поскольку число генов в организме несоизмеримо больше числа хромосом, то понятно, что каждая хромосома несет много генов. Каждый ген занимает в хромосоме определенное место – локус. Гены, расположенные на одной хромосоме, называются сцепленными.
Помимо ядра, небольшая доля генетической информации эукариотической клетки находится в таких органоидах, как митохондрии и хлоропласты, имеющих собственные генетические системы: свои ДНК, разнообразные РНК (и-РНК, т-РНК, р-РНК) и рибосомы, что позволяет осуществлять независимый синтез белка. Кольцевые ДНК этих органоидов явились важным доводом в пользу их бактериального симбиотического происхождения на заре формирования жизни.
Клеточное ядро эукариот разделяет процессы транскрипции и трансляции, что предоставляет широкие возможности для регуляции. Регуляция происходит на всех этапах экспрессии генов эукариот. Дополнительным этапом у них является процессинг – процесс сложных преобразований синтезированной в ходе транскрипции РНК. Важнейшая составляющая процессинга и-РНК – сплайсинг, при котором происходит вырезание интронов (некодирующих областей гена) и сшивание экзонов (кодирующих областей). Экзоны и интроны обусловливают «мозаичную» структуру эукариотических генов. Именно в результате процессинга синтезированная в ядре РНК становится функционально активной.
Понимание многообразных механизмов регуляции послужило причиной радикальных изменений наших представлений о структурно-функциональной организации генетического аппарата в настоящее время.
Одним из основателей современной генетики, выдающимся датским ученым В. Иоганнсеном (1857–1927) были предложены базовые генетические термины – ген, аллель, генотип, фенотип, определяющие генетические характеристики особи.
Гены, расположенные в своих локусах, могут иметь варианты – аллели. Локус, имеющий в популяции более одного аллеля, называется полиморфным. Обычно аллели обозначаются буквами латинского или греческого алфавита, а если их много, то с верхним индексом. Количество аллелей разных генов в популяциях организмов может быть различным. Одни гены имеют много аллелей, другие – мало. В любом случае число аллелей ограничено эволюционными факторами: аллели, ухудшающие адаптивные свойства вида либо несовместимые с жизнью, элиминируются естественным отбором.
Конкретный организм эукариот имеет только два аллеля одного гена: по числу гомологичных локусов гомологичных хромосом (отцовской и материнской). Организм, у которого оба аллеля одинаковые, называется гомозиготой (по данному гену). Организм, у которого аллели разные, называется гетерозиготой (рис. 1.4). Аллели, локализованные на половых хромосомах гетерогаметного пола, могут присутствовать в единственном числе.
Генотип можно представить как совокупность аллелей организма, а фенотип – как совокупность его внешних признаков.
Введенный в 1920 г. немецким ботаником Г. Винклером (1877–1945) термин геном стал характеристикой целого вида организмов, а не конкретной особи. Это понятие в дальнейшем стало одним из важнейших. К 1980-м гг. XX в. формируется новое направление генетики – геномика. Первоначально геном характеризовали как совокупность генных локусов гаплоидного набора. Однако оказалось, что сами гены занимают относительно небольшую часть генома, хотя и составляют его основу. Большую часть занимают межгенные участки, где есть области с регуляторной функцией, а также районы невыясненного пока назначения. Регуляторные участки неразрывно связаны с генами, являются своего рода «инструкциями», определяющими работу генов на разных этапах развития организма. Поэтому геномом в настоящее время называют всю совокупность ДНК клетки, характерную для ДНК вида.
На современном этапе развития генетики геномика становится одним из ее ключевых разделов. Успехи геномики наглядно продемонстрировало успешное завершение программы «Геном человека».

Рис. 1.4. Аллели сцепленных генов двух гомологичных хромосом
1.3. Проявление генетической информации
Как проявляется генетическая информация, локализованная в ДНК-содержащих структурах? Ее проявление отражается в фенотипе, который, согласно современным представлениям, всегда является результатом взаимодействия генов в конкретных условиях внешней среды.
В природе обнаружены самые разные варианты взаимодействия аллелей и неаллельных генов. Ген может подавлять или видоизменять проявление другого гена (Курчанов Н. А., 2009; 2010). На результат такого взаимодействия влияет множество факторов, поэтому перед генетиками открывается все более и более сложная картина. Сами гены стали рассматривать не как единичные образования, а как составляющие единой структуры – транскрипционной сети. «Включение» каждого гена сети производится сигнальными молекулами, причем взаимоотношения между генами внутри сети могут быть очень сложными. В результате этого признак может проявляться (или не проявляться) у особей одного генотипа, а степень проявления признака может варьировать в различных условиях среды.
Проявляемость гена в фенотипе определяет такой показатель, как пенетрантность, степень его выраженности – экспрессивность, а диапазон варьирования признака (проявлений генотипа) – норма реакции. Этот диапазон различен для разных признаков. Некоторые признаки имеют однозначную норму реакции или варьируют незначительно. Другие признаки (например, поведенческие) имеют широкую норму реакции.
Наряду с наследственностью, всем живым организмам свойственна изменчивость, под которой понимают свойство приобретать новые признаки. В природе встречаются различные виды изменчивости.
Мутации – это наследственные изменения генетического материала. По характеру изменений генома мутации делятся на генные (изменения нуклеотидного состава ДНК отдельных генов), хромосомные (изменения структуры хромосом), геномные (изменения числа хромосом), цитоплазматические (изменения неядерных генов). Эти основные группы мутаций имеют множество подгрупп, которые подробно рассматриваются в курсе генетики (Курчанов Н. А., 2009; 2010).
Несмотря на неблагоприятный эффект большинства мутаций, способность к мутированию чрезвычайно важна, так как дает возможность организмам изменяться и приспосабливаться к условиям внешней среды. Мутации – основа эволюционных изменений.
Модификации – это изменения фенотипа под действием факторов внешней среды без изменения генотипа. Они обеспечивают приспособленность организма в течение жизни к условиям его существования. Проблема наследования модификаций имеет долгую историю чрезвычайно острых дискуссий. Не меньший интерес представляет проблема возникновения модификаций. Способность к адекватным модификационным изменениям открывает организму более широкие возможности для адаптаций.
Дискуссии о возможностях модификаций особенно болезненно проходили в сфере поведенческих наук. Поведенческие признаки высших организмов контролируются большим числом генов, что определяет широкий диапазон фенотипической изменчивости. Поведенческие признаки, контролируемые единичным геном, могут быть предметом строгого генетического анализа, но только у простых организмов. В основном такие признаки исследованы у дрозофилы – классического объекта генетики. Для выражения доли изменчивости, обусловленной различиями в генотипе, был введен показатель наследуемости.
Однако оказалось, что понятие наследственности не исчерпывается генотипом. Один и тот же генотип может по-разному «работать», а «режим» его «работы» может передаваться по наследству. На волне революционных открытий конца XX в. рождается новое направление – эпигенетика.
1.4. Эпигенетика – новый взгляд на природу наследственности
Эпигенетика в настоящее время «поставляет» наиболее интересные факты для «строительства» новой парадигмы современной генетики. Термин «эпигенетика» был предложен известным английским генетиком К. Уодингтоном (1905–1975) еще в 1942 г. для обозначения процессов формирования конкретного фенотипа на основе определенного генотипа. Первоначально под такими процессами подразумевались обратимые модификации в ходе онтогенеза.
В настоящее время особое внимание уделяется наследственным изменениям генной экспрессии, происходящим без изменения нуклеотидного состава ДНК. Фундаментальным положением эпигенетики является то, что один и тот же генотип может проявляться разными фенотипами, которые опосредуются разными состояниями генных локусов. Для выражения этих состояний, обусловленных различными эпигенетическими механизмами, появился особый термин – эпигеном (Паткин Е. Л., 2008). Выделяют несколько эпигенетических механизмов.
Метилирование ДНК – наиболее распространенный и изученный вариант включения и выключения генной экспрессии при помощи метильной группы, связанной с азотистым основанием (обычно цитозином). Метилирование осуществляется целым семейством ферментов метилтрансфераз. Особенно велика его роль на ранних этапах развития, когда протекают процессы дифференцировки.
Изменение структуры хроматина в ходе онтогенеза имеет не меньшее значение. Основной способ такой регуляции – разнообразные модификации гистоновых белков нуклеосом. С этими модификациями связана активация или выключение специфичных генов. Модификации гистонов играют ключевую роль в формировании гетерохроматина (конденсированного нефункционирующего хроматина). Кроме гистонов, в хроматине содержится большое количество других белков, получивших собирательное название негистоновых. Модификации таких белков также влияют на активность генов.
Воздействие малых РНК оказалось сопоставимо по своей значимости для регуляции генной экспрессии с активностью регуляторных генов. Многие малые РНК происходят из межгенных участков и интронов матричной РНК, вырезаемых в процессе сплайсинга. Их многообразие обусловливает наличие различных классификаций малых РНК.
РНК-зависимый механизм регуляции экспрессии генов, открытый только в 1995 г., получил название РНК-интерференции. В основном он запускается при помощи двунитевых малых РНК (Смирнов А. Ф., 2009). Это открытие радикально изменило наши представления о регуляции генной экспрессии в ходе онтогенеза.
Влиянием малых РНК, может быть, объясняется загадочное явление парамутации, при котором аллель «запоминает» свое состояние в предыдущем поколении. Это явление противоречит законам Менделя, согласно которым гены гетерозигот расходятся в половые клетки неизменные. Было предложено несколько моделей, объясняющих возникновение парамутаций, но все они не более чем гипотезы. Возможно, новому поколению при оплодотворении передаются молекулы малых РНК. Вероятно, парамутации широко распространены в природе, обусловливая явления пенетрантности и экспрессивности (Паткин Е. Л., 2008).
Наиболее ярким эпигенетическим феноменом является геномный импринтинг, открытый в 1984 г. Он проявляется у млекопитающих и цветковых растений. Геномный импринтинг представляет собой обусловленность экспрессии генов в зависимости от отцовского или материнского происхождения. Зигота, а затем и клетки организма как бы «запоминают» происхождение гена. Несмотря на интенсивное изучение в последние годы, механизм геномного импринтинга во многом остается непонятным. Вероятно, он обусловлен различием в метилировании хромосом.
Предположительно, 0,1 % генов млекопитающих подвергаются импринтингу (Смирнов А. Ф., 2009). Хотя число импринтированных генов относительно невелико, все они играют важную роль, особенно в регуляции поведения. Именно из-за наличия импринтированных генов необходимо наличие отцовского и материнского геномов для нормального развития зародыша.
В исследованиях генетиков рассматриваются и другие эпигенетические механизмы регуляции генной экспрессии. Они, безусловно, играют свою роль в развитии большинства полигенных болезней человека (Cook G., Hill A., 2001). Относительно недавно показана важная роль влияния внутриутробного развития на организм и его последующее поведение. Так, сходство разнояйцовых близнецов значительно больше, чем сходство обычных братьев и сестер, хотя степень генетической общности в обоих случаях одинакова (Scarr S., 1992).
Дальнейшее изучение эпигенетических механизмов должно раскрыть такое явление, как структурная наследственность, под которой понимается функционирование в качестве матриц трехмерных структур, образующих себе подобных. Самым известным примером являются белки прионы (Шаталкин А. И., 2009).
Более пристальное внимание к эпигенетическим механизмам раскрыло возможность внешнего воздействия на наследственность. Даже такие внешние факторы, как питание, материнская забота, способ оплодотворения, влияют на эпигеном (Паткин Е. Л., 2008). Это влияние может осуществляться путем биохимической модификации ДНК и гистонов, что изменяет генную экспрессию. Если такие модификации затрагивают линии первичных половых клеток (ППК), то происходит наследование фенотипа. Показана возможность передачи по наследству «рисунка метилирования» генов, что изменяет фенотипические свойства.
Таким образом, некоторые эпигенетические феномены бросают вызов традиционной генетической парадигме. Вековая проблема «наследования приобретенных признаков» возвращается в генетику в новом качестве.
Автономность генов оказалась значительно меньшей, чем предполагалось на заре генетики. Именно поэтому так трудно дать четкое определение гена. Гены генома интегрированы в транскрипционные сети, которые можно рассматривать как самостоятельные структуры генетического аппарата. Их взаимосвязь открывает возможности моделировать и передавать по наследству специфическую активность генома.
А. И. Шаталкин приводит интересное сравнение деятельности организма и современного производства. При этом генотип можно сравнить с технической документацией, а фенотип – с конечным продуктом. Но на производстве нельзя не учитывать роль рабочего коллектива, который реализует технический проект. А в организме? Возможно, эту роль выполняют процессы самоорганизации (Шаталкин А. И., 2009).
Впервые термин «самоорганизующаяся система» был использован одним из основателей кибернетики У. Р. Эшби (1915–1985) еще в 1947 г. (Ashby R., 1956). В настоящее время вопросы самоорганизации активно разрабатываются в рамках синергетики, чье влияние на биологию все более ощутимо. Такой синтез заставляет ученых по-новому понимать явления наследственности и изменчивости, служит веским фактором разработки новой парадигмы.
Другим весомым фактором послужили открытия, опровергающие наши представления о стабильности генома.
1.5. Лабильный геном
Традиционные представления о стабильности геномов, сложившиеся в рамках классической генетики, сильно пошатнулись после открытия мобильных (мигрирующих) генетических элементов (МГЭ). МГЭ – это структуры, которые могут перемещаться в пределах генома или переходить из генома в геном. Они могут встраиваться в различные области генетического аппарата эукариот и прокариот. Процесс перемещения МГЭ получил название транспозиции.
МГЭ были открыты Б. Мак-Клинток (1902–1992) у кукурузы (Zea mays) еще в 1940-е гг., однако ее сообщение было встречено генетиками с большим недоверием. Развитие молекулярной генетики подтвердило наличие подобных структур сначала у прокариот, а затем и у эукариотических организмов. Только в 1983 г. Б. Мак-Клинток была удостоена Нобелевской премии за свое выдающееся открытие.
В настоящее время ко всем МГЭ обычно применяют термин транспозоны.
Транспозоны были обнаружены у бактерий, дрожжей, кукурузы, дрозофилы и других эукариотических организмов. Типичные признаки транспозонов – инвертированные повторы на концах и дупликация сайт-мишени (места внедрения). Развитие методов молекулярной генетики показало широкое распространение транспозонов в геномах высших животных, в том числе и у человека. Их доля в геномах оказалась значительно выше, чем предполагалось ранее. Так, в геноме человека транспозоны составляют до 45 % генетического материала (Смирнов А. Ф., 2009).
В своем большинстве транспозоны являются эволюционным наследием и обусловлены внедрением вирусного или бактериального генома в клетки эукариот миллионы лет назад. Особый интерес представляют ретровирусы, которые реализуют путь, альтернативный «центральной догме». Это феномен обратной транскрипции – способность строить на своей РНК комплементарную ДНК. Такая ДНК-копия, встраиваемая в чужой геном, получила название провируса.
Подавляющее большинство провирусов не функционируют (не экспрессируются), но при определенных воздействиях некоторые из них способны активироваться, что может представлять собой опасность для макроорганизма. Однако некоторые процессы развития, например развитие нейронов, закономерно сопровождаются повышенной активностью транспозонов. Вероятно, явление ретротранспозиции сыграло важную роль в эволюции эукариот. У человека ретротранспозоны составляют более 90 % мобильной части генома (Попов В. В., 2008).
Феномен транспозиции представляет большой теоретический интерес, независимо от конкретной функции МГЭ. Их важная роль в эволюции уже не подлежит сомнению: показана близость сайтов внедрения МГЭ и хромосомных перестроек. Поскольку МГЭ могут создавать диффузные области гомологии, их копии в разных местах генома обеспечивают возможность реципрокной сайт-специфичной рекомбинации. Такие обмены могут приводить к различным хромосомным перестройкам (делециям, инверсиям, транслокациям), могут изменять систему регуляции генов. Стимулируя хромосомные перестройки, транспозоны могли изменять экспрессию соседних генов. Такая реорганизация играет существенную роль в эволюции геномов.
Показан резкий рост числа транспозиций в популяциях дрозофилы при стрессовых воздействиях, что можно рассматривать как модель быстрой регуляции активности генома при изменении условий существования (Ратнер В. А., 2002).
В настоящее время наметилась тенденция оставить понятие «мутация» только для генных мутаций, а хромосомные и геномные мутации рассматривать как варианты естественной генетической рекомбинации. Терминологическая проблема возникла из-за традиционного понимания мутации как отклонения от нормы. Однако дупликации, инверсии, транслокации, центрические слияния, полиплоидия неразрывно связаны с эволюцией геномов. С эволюционной точки зрения, хромосома – это непостоянная структура, а лабильность – естественное состояние генома (Хесин Р. Б., 1984).
Все вирусы представляют собой «мобильные гены». Вирусы, плазмиды (мелкие кольцевые ДНК у бактерий), транспозоны можно рассматривать в русле единого явления в эволюции генетической системы. Все эти структуры не только сами способны перемещаться по геному или из генома в геном, но и могут «перетаскивать» соседние участки генома клетки-хозяина. Такое явление получило название трансдукции. Его открытие сыграло решающую роль в рождении генной инженерии.
Многообразие мобильных генетических структур послужило основой для формирования гипотезы горизонтального переноса генов в эволюции. Этим термином стали называть обмен генетической информацией между неродственными организмами неполовым путем. Хотя участие транспозонов в процессе эволюции не отрицает никто, их значение до сих пор вызывает бурные споры. Одни генетики, допуская возможность горизонтального переноса в природе, не считают, что он играл заметную роль в эволюции. Другие, наоборот, придают ему решающее значение.
Мы не можем пока адекватно оценить эволюционные масштабы горизонтального переноса, но генетические исследования все чаще показывают его распространенность в природе. Уже сейчас ясно, что роль горизонтального переноса важнее, чем предполагалось ранее. Особенно значимо это было на первых этапах эволюции, когда происходило формирование основных групп живых существ.
У высокоорганизованных организмов генетическую рекомбинацию обеспечивает половой процесс, что дает необходимую им стабильность. Но нельзя игнорировать роль горизонтального переноса и для высших организмов. Так, многие важные явления, связанные с иммунитетом зародышей млекопитающих, вероятно, находятся под контролем генов, полученных от вирусного генома путем трансдукции. Выдают свое происхождение от транспозонов и многие регуляторные последовательности геномов позвоночных (Марков А., 2010).
Возможно, генотипы всех организмов представляют собой «генофонд мира» как единую структуру (Попов В. В., 2008). Этот взгляд не лишен оснований, что заставляет нас с осторожностью относиться к филогенетическим построениям с помощью геносистематики, по-новому посмотреть на многие положения эволюционной теории, с которой мы познакомимся в следующей главе.
