Часть IV. Кайнозой: ещё не вечер?
Палеоген. 66–23,03 миллиона лет назад: от мира карликов до мира гигантов
Международная и российская шкала:
66 млн л. н. – палеоцен: датский век – 61,6 – зеландский век – 59,2 – танетский век – 56 – эоцен: ипрский век – 47,8 – лютетский век – 41,2 – бартонский век – 37,71 – приабонский век – 33,9 – олигоцен: рюпельский век – 27,82 – хаттский век – 23,03
Палеоцен. 66–56 миллионов лет назад: мир карликов
Первый период кайнозойской эры – палеоген, а его первая эпоха – палеоцен. Гигантские ящеры вымерли, а на их место заступили карлики – крошечные наследники мезозоя, сначала робко, но со всё большим энтузиазмом начавшие реставрировать пострадавшую от вымирания биосферу. Среди них совсем немаловажное место занимали наши прямые предки – первые приматоморфы.
Карта начала палеогена похожа и не похожа на современную. Без труда узнаются все континенты и архипелаги, океаны и заливы, но во всём есть какая-то странность. Европа – остров, отделённый от Азии широким морем-проливом, в окружении россыпи мелких островков. Южная Азия – вытянутый по широте архипелаг, на дне проливов которого кроются будущие вершины Гималаев. Индия и вовсе дрейфует посреди будущего Индийского океана. Аравия ещё не отделилась от Африки и никак не соединена с Азией, а поперёк Сахары досыхает огромный залив Средиземного моря. Между зарождающейся Индонезией и Австралией, только что отпочковавшейся от Антарктиды, лежит огромное море – слияние Индийского и Тихого океанов. Атлантический океан почти замкнут на севере и совсем узенький на юге, зато на месте Панамы вольготно и плавно перетекает в Тихий, а поднимающиеся Анды отделены от Амазонии длинным узким заливом Карибского моря.
Весь предыдущий меловой период на дне морей откладывался CaCO3, то есть мел, собственно, и давший последнему периоду мезозоя его название. Но время шло, тектоника неумолимо ворочала литосферные плиты, и местами – в зонах океанических разломов – рано или поздно донные отложения погружались в глубины планеты (для этого есть умный термин «субдукция»). Безграничные толщи мела плавились в магме, отчего CO2 высвобождался и поднимался в атмосферу, вызывая парниковый эффект и приводя к потеплению в общепланетарных масштабах. Так в отдалённой перспективе сработало наследие водорослей кокколитофорид, когда-то снизивших парниковый эффект и тем заморозивших динозавров. Ныне же, когда злые ящеры сгинули, температуры вновь скакнули вверх. Правда, на протяжении палеоцена эта тенденция срабатывала ещё не вполне однозначно, и пару раз даже намечалось похолодание, но в любом случае климат был гораздо теплее большей части и мелового, и юрского, а уж по сравнению с нынешним и подавно – аж на 10 °C курортнее! В целом жизнь однозначно налаживалась!
Местами это восстановление можно проследить очень детально. Пепелища пожаров, вызванных падением метеорита, густо поросли грибами – пресловутый иридиевый слой, отложившийся при оседании пыли от падения астероида, просто перенасыщен грибными спорами (между прочим, подобная же история намного раньше случилась на границе перми и триаса, где грибы тоже первыми встречали первые лучи мезозоя на руинах палеозоя). Кстати, часть спор могут быть и не грибными, а водорослевыми, что тоже логично: симбиоз грибов с водорослями – лишайники – универсальные выживальщики, первыми заселяющие любую пустошь, так как водорослевый фикобионт способен к фотосинтезу, то есть берёт энергию из света, а из воды и углекислого газа синтезирует органику, тогда как грибной микобионт разлагает органику обратно. Никакие консументы – существа, перерабатывающие органику в другую органику, то есть мы, – в таком раскладе вовсе не нужны, они лишни и избыточны. Через некоторое время эти грибные плантации начали снова зарастать лесами.
В колорадском Коррал Блюффс палеонтологи крайне точно датировали подробнейшую последовательность напластований с массой ископаемой флоры и фауны. Оказывается, что непосредственно после мел-палеогеновой катастрофы резко увеличилось количество папоротников, пальм, за следующие сто тысяч лет разнообразие млекопитающих удвоилось, а за триста тысяч лет размеры их тел утроились. Через семьсот тысяч лет после границы колорадщину топтали уже очень приличного размера звери, например полусоткилограммовый Eoconodon coryphaeus, отъедавшийся на свежепоявившихся бобовых. Крокодилы и черепахи дополняли картину тропического болота. Но эти снулые рептилии уже не могли остановить торжества пушистых и теплокровных зверей!
Современные бобовые растения Fabacea – чудесные создания! Во-первых, у них шикарные двусторонне-симметричные цветы с широкими лепестком-парусом сверху, узкими вёслами по бокам и парой соединённых в одну лодочку – снизу. Когда пчёлка или другой какой опылитель ползёт к желанному нектару по такой конструкции, на тельце гарантированно налипает пыльца. Для этого нижние девять тычинок обычно срастаются в желобок и намазывают пыльцу на пузико пчёлке, а одна свободная тычинка аккуратно, но неумолимо шлёпает сверху, прилепляя пыльцу на голову и спину. Опыление при таком раскладе просто неизбежно!
Вторая, куда более важная особенность бобовых – клубеньки на корнях, домики для клубеньковых азотфиксирующих бактерий Rhizobiales. Эти бактерии выбирают из атмосферы несвязанный азот и заключают его в состав органических молекул. А это процесс не такой простой. Азота-то в атмосфере у нас навалом, а вот связывать его умеют немногие, при том, что азот входит в состав кучи необходимых всем органических соединений, включая нуклеиновые кислоты. Наличие бобовых на любом луге несказанно увеличивает плодородие почвы, так как все прочие растения тоже могут получить связанный азот на дармовщинку.
Появление бобовых неизбежно должно было мощно двинуть эволюцию растительных экосистем. Оно и двинуло, планета начала зарастать густыми лесами – нашим домом на долгие последующие миллионы лет.
Морские экосистемы кайнозоя восстанавливались едва ли не быстрее наземных. Сильнее всего поменялись самые нижние и самые верхние экологические уровни – планктон и хищники. Полностью исчезли все хищные морские рептилии – плезиозавры и мозазавры. Шанс на захват океана получили хрящевые рыбы – химеры, акулы и скаты. Любопытно, что в конце мела сгинули лишь две их большие группы – Hybodontida и Synechodontiformes, причём вторые, во-первых, всё же обнаруживаются в палеоцене, а во-вторых, могли даже не вымереть, а стать предками для некоторых современных хрящевых. Однако недостатки их дыхания – отсутствие жаберных крышек – и системы поддержания равновесия – отсутствие плавательного пузыря – видимо, уже были неисправимы. Конечно, и сейчас повстречаться с акулой в открытом море – сомнительная радость (вернее, радость, но только для акулы), а несколько позже – в эоцене и далее – они даже дали новое разнообразие, но свой шанс хрящевые так до конца и не реализовали.
Без особых проблем пережили катаклизм осетры, а вот костные ганоиды сохранились лишь в виде реликтов. Из позднего мела в палеоген и вплоть до современности переходят роды осётров Acipenser и панцирных щук Lepisosteus, с палеоцена известны и амии Amia.
На этом безрыбье костистые рыбы вдруг оказались полновластными царицами морей. Мы ведь сейчас живём на планете рыб – число их видов и жизненных форм намного больше, чем всех прочих позвоночных, вместе взятых.
Конечно, и им позднемеловое вымирание аукнулось более чем основательно. Огромное число линий пресеклось, а после из остатков быстро заново сформировались новые формы. Частично они повторяли то, что уже плавало в мезозое.
Из всего этого великолепия особое внимание обращает на себя Serrasalmimus secans из палеоцена Марокко. Его родичи были весьма колоритны и более разнообразны до вымирания, но и выживший потомок не подвёл: его челюсти были плотно уставлены страшными треугольными зубами, практически как у современных пираний. Скорее всего, сходство это не обусловлено родством, а конвергентное, то есть объясняется приспособлением к одинаковым условиям.
Без особых проблем преодолели вымирание амфибии, черепахи и крокодилы. Те же роды, тот же темп исчезновения и появления новых видов: астрономия и геология, все эти ваши астероиды и вулканы, этих существ явно не волновали.
Хуже пришлось ящерицам и змеям: в фаунах Северной Америки исчезло пять шестых видов; правда, тут же несколько и появилось. Показательно, что пропали самые крупные твари – весом более полукилограмма. Значимое разнообразие чешуйчатые восстановили уже в благословенном эоцене. Впрочем, размеры росли быстрее изменчивости: в конце палеоцена колумбийский гигантский удав Titanoboa cerrejonensis достиг 13 м и веса более тонны. Это круче даже многих мезозойских ящеров. Позвонки современных удавов – отнюдь не маленьких, трёх с половиной метровых – выглядят на фоне позвонков титанобоа просто крошечными. Такие монстры явно должны были и питаться кем-то соответствующим. Сопутствующая фауна включает двоякодышащих и костистых рыб, черепах и специфических крокодилов Dyrosauridae. Последние – Anthracosuchus balrogus, Acherontisuchus guajiraensis и Cerrejonisuchus improcerus, – видимо, были главной добычей титанической змеи. Между прочим, здешняя черепаха Carbonemys cofrinii – тоже рекордсменка: её панцирь был 1,72 м в длину! Что там было подмешано в воду, что их так пёрло?! Весь этот серпентарий бултыхался в жарких тропических болотах Южной Америки под покровом протосельвы – прототипа нынешних джунглей. Млекопитающих там, кстати, не замечено, хотя в принципе на материке они в это время были. Получается, век гигантских рептилий в Южной Америке с концом мезозоя вовсе не закончился! И лишь спустя некоторое время звери и тут взяли своё.

Titanoboa cerrejonensis
Как часто бывает, исчезновение одних привело к появлению других: среди чешуйчатых рептилий возникли амфисбены Amphisbaenia. Древнейшими представителями были Oligodontosaurus wyomingensis, Archaerhineura mephitis и Chthonophis subterraneus из Вайоминга, а также Polyodontobaena belgica и Camptognathosaurus parisiensis из Бельгии и Франции. Их челюсти, плотно усаженные толстыми зубами, очень уж характерны.
Современные амфисбены Amphisbaenia чаще рассматриваются как подотряд чешуйчатых, но иногда предлагается выделить их в самостоятельный отряд, настолько они своеобразны. В отличие от ящериц и змей, тело амфисбен покрыто сплошной роговой оболочкой, подразделённой поперёк и немножко вдоль, отчего внешне амфисбены почти неотличимы от дождевых червей. Кожа слабо соединена с телом, так что легко сдвигается, позволяя амфисбенам двигаться едва ли не перистальтически, причём как вперёд, так и назад, что, во-первых, объясняет альтернативное название «двуходки», а во-вторых, ещё больше делает их похожими на червей.
Но достаточно посмотреть на череп, чтобы стало ясно: амфисбены – вовсе не мирные червячки! Череп амфисбен максимально похож на череп «чужого» из известного фильма: вытянутый, массивный, почти безглазый, со страшными зубастыми челюстями (что довольно странно, учитывая питание термитами, муравьями и дождевыми червями; впрочем, вспоминая кротов и землероек, может, не так уж и странно), а вместе с формой тела – на червей из «Дюны»[1]. Головой амфисбены копают, так что форма заточена под это дело, причём у разных родов и видов реализованы вариации роющих инструментов: под плоскую лопату, под заострённую мотыгу, под кол или отбойный молоток, причём рабочий передний край может быть опущен или курносо поднят. Амфисбены могут ползать и по-змеиному, и изгибаться по вертикали, а также поднимать гибкий хвост, не отличающийся от головы, для отвлечения внимания, что, кстати, даёт их очередное название – «двухголовые змеи».
Почвенный образ жизни диктует и другие особенности. Глазки у амфисбен редуцированы и скрыты под кожей, откуда едва просвечивают. Сохранилось только левое лёгкое, кстати, в отличие от змей и безногих ящериц с их одним правым, что заодно показывает независимость эволюции амфисбен. Ручки и ножки пропали, и только североамериканские Bipes сохранили крошечные передние ручки – трёх- или пятипалые.
Загадочно географическое распространение амфисбен: возникли они в Северной Америке, откуда мигрировали в Европу и Африку, а уже из Африки – сильно позже и причём дважды независимо – обратно через Атлантику – один раз прямо на Карибы и второй – туда же, но транзитом через Южную Америку. Выходит, амфисбены несколько раз успешно преодолели морские просторы, будучи почвенными роющими созданиями! После этого остаётся лишь удивляться, как же малоподвижны были наши предки!
На пернатых катаклизм сказался странно-избирательно: часть линий типа энанциорнисов и конфуциусорнисов исчезла без следа, а веерохвостые птички сдюжили. Как обычно с хрупкими пташками, при изучении древних фаун надо делать основательную скидку на плохую сохранность эфемерных косточек.
Как минимум на границе палеоцена и эоцена уже существовал отряд древненёбных птиц-палеогнат Lithornithiformes. Эти птички, например много видов Lithornis, в наибольшей степени (и внешне, и по сути) напоминали современных южноамериканских тинаму или (не по родству, а чисто внешне) – куропаток. Строение их нёба выдаёт принадлежность к древненёбным птицам, большой киль явно свидетельствует о хороших летательных способностях, а длинные ноги намекают на околоводность. Судя по мозгу Lithornis plebius, эти птицы имели отличное обоняние, хотя и уступающее современным киви, но сопоставимое с тинаму. Благодаря шикарной сохранности черепа Lithornis promiscuus, а также некоторых других родственных птиц, удалось установить, что кончик клюва литорнисов был очень чувствительным, как у современных тинаму, киви и ибисов. Такая вибротактильность, очевидно, нужна птицам, постоянно ковыряющимся в земле, так что легко представить берега североамериканских и европейских болот раннего кайнозоя, по которым среди травы снуют «курочки» и тычут длинными острыми клювиками в тину.
Современные тинаму Tinamiformes – внешне невзрачные, но очень важные для эволюции птички. Они весьма похожи на куропаток, однако родственны южноамериканским страусам нанду, что особенно здорово видно по поведению. У тинаму неплохое по птичьим меркам обоняние; питаются они всем подряд. Тинаму имеют большой киль на грудине, но их грудная мускулатура белая, то есть имеет мало митохондрий и не может обеспечить долгий надёжный полёт. Поэтому тинаму предпочитают прятаться и застывать на месте, лишь в крайнем случае убегая и уж совсем в безнадёжном – взлетая. Но их коротенький хвостик не всегда позволяет лететь куда полагается, так что тинаму могут даже врезаться в коварные деревья, бросающиеся наперерез. Гнёзда у тинаму очень простенькие, а многие виды откладывают яйца просто на землю. Как и у страусов, в одно гнездо откладывают яйца несколько самок, а высиживает самец; он же потом печётся о птенчиках, которые, впрочем, весьма самостоятельны с момента вылупления. Тинаму очень уязвимы, почти все птенцы гибнут, зато и взрослеют меньше чем за месяц. Все эти особенности прямо отсылают к дремучим мезозойским временам. Неспроста тинаму сохранились лишь в Южной Америке, где бодрых плацентарных хищников не так много. Впрочем, там их не так уж мало – с полсотни видов.
Древненёбные птицы с самого начала были склонны к наземной жизни и быстро теряли способность к полёту, а уж с исчезновением хищных динозавров эта тенденция получила отличные возможности для реализации. Тут же появился целый веер нелетающих страусоподобных птиц: например, Remiornis heberti из Франции и Diogenornis fragilis из Бразилии – птицы неясного родства, но, вероятно, близкие страусам-нанду.
Древненёбные птицы, впрочем, не задались. На первое место выпорхнули новонёбные.
Ясно, что наилучшим образом известны околоводные аналоги или даже прямые родственники гусей и уток. Несколько видов Presbyornis на своих колченогих лапах вышагивали по камышам Северной Америки и Центральной Азии, вытягивали длинные шеи и собирали широким вогнутым клювом – прямиком из «Утиных историй» – всяческую тину.
Гусеобразные птицы жили и в Южном полушарии: почти полная внешняя копия пресбиорниса Conflicto antarcticus летала над самой Антарктидой, которая тогда имела вполне умеренный климат и была покрыта лесами, а Australornis isoni из Новой Зеландии был родственен меловым антарктическим Vegavis iaai и Polarornis gregorii.
В той же Новой Зеландии обнаружен и самый примитивный и одновременно самый маленький пелагорнис Protodontopteryx ruthae. Пелагорнисы Pelagornithidae – странная группа птиц, появившаяся в начале кайнозоя и вымершая совсем недавно – когда наши предки уже начинали пользоваться орудиями труда. Две черты пелагорнисов обращают на себя особое внимание. Во-первых, это были похожие на альбатросов планёры от полутора до шести метров в размахе крыльев – одни из самых огромных птиц за всю историю планеты. Во-вторых, их челюсти были какой-то зверской пилой, так как их края покрывали далеко отстоящие, но очень большие зазубрины. Это не были настоящие зубы, а именно выросты челюсти; поэтому альтернативное название пелагорнисов – «псевдозубатые птицы». Вероятно, такими клювами удобно было выхватывать из воды кальмаров и крупных, но не слишком бронированных рыб. Скорее всего, первоначальный успех пелагорнисов был связан с вымиранием гигантских морских птерозавров, а также зубатых птиц ихтиорнисов. Экологическая ниша планеров-рыбоядов пуста не бывает! Видимо, и итоговое вымирание было связано с изменениями в рыболовецкой области, скорее всего – с появлением крутых конкурентов типа современных альбатросов и чаек. Впрочем, довольно долго пелагорнисы сосуществовали с современными птицами-рыбоядами, так что конкурентная гипотеза явно не самодостаточна. Учитывая время исчезновения – 2,5 миллиона лет, то есть время мощных перестроек климата, – причина, скорее всего, хитро-экологическая, но детали процесса пока ускользают. Впрочем, это было сильно позже палеоцена…
Понятно, что рыбу можно ловить по-разному. Исчезли же не только летуны – птерозавры и ихтиорнисы, но и нелетающие гесперорнисы. Экологическую нишу прибрежных ныряльщиков тут же, конечно, заняли пингвины Sphenisciformes. Богатейший палеоценовый пингвинятник открыт в Новой Зеландии: древнейший и примитивнейший Muriwaimanu tuatahi, его чуть более поздняя и продвинутая версия Waimanu manneringi, ещё более прогрессивные Kumimanu biceae и Crossvallia waiparensis, а также иные глупые птицы робко прятали тела жирные в утёсах. Уже муривайману и вайману были размером с императорского пингвина, кроссваллия достигала роста 160 см и веса 70–80 кг, а кумиману – 160–180 см и 90–100 кг! Даже самые ранние пингвины были вполне узнаваемы: мешковидное вертикальное тело на коротких ножках, загребущие крылья-ласты, длинный острый клюв для ловли рыбы. Довольно быстро они распространились по Южному полушарию, где и процветают поныне.
Имелись птицы и попроще. Над мелководными лиманами моря Волгоградской области порхали Volgavis marina, похожие на серебристых чаек, но, возможно, более родственные фрегатам или буревестникам.
Первоначально волгавис был датирован поздним маастрихтом, но после возраст оказался раннепалеоценовым. Кстати, в одной статье с описанием волгависа упоминается плечевая кость птерозавра-аждархида из недалёкого от волгависа местонахождения, найденный в позднепалеоценовом слое и признанный перемытым из маастрихтского. Но, строго говоря, переотложенность исходит скорее из самого факта птерозавровости. А так-то, почему бы отдельным птерозаврам и не пожить чуток в палеоцене?.. Фантазия, конечно, но, возможно, стоило бы обращать на подобные находки особое внимание?
Длинноногая птица Scaniornis lundgreni, вероятно, похожая на фламинго (что, впрочем, весьма спорно, так как остатки крайне фрагментарны), шагала по берегам палеоценовых Швеции и Германии. Целая орнитофауна границы мела и палеоцена обнаружена в формации Хорнестаун в Нью-Джерси: гусь Anatalavis rex, буревестник Tytthostonyx glauconiticus и фаэтон Novacaesareala hungerfordi. Как и в предыдущих случаях, тут найдена околоводная фауна. Что происходило в лесах и горах – большей частью покрыто мраком.
Впрочем, и про это немножко известно благодаря находкам во Франции, в местонахождении Серне-ле-Реймс и некоторых других. Самыми эффектными палеоценовыми лесными птицами были, без сомнения, Gastornis parisiensis и G. russeli – ближайшие родственники кур. Эти двухметровые нелетающие «петушки» имели толстенные трёхпалые ноги и огромадный клюв, похожий на клещи-болторезы для срезания стальных цепей и штырей. Конечно, такой колоритный облик был интерпретирован как хищническая адаптация: якобы гасторнисы были злобными притеснителями несчастных первобытных зверюшек. Образ демонической убер-курицы, терзающей панически разбегающихся протолошадок, прочно вошёл в палеогеновый бестиарий популярных книг о палеонтологии. Однако новейшие исследования показывают, что реальность, видимо, была совсем не такой. Изотопные анализы и изучение следов прикрепления мышц на челюстях показали, что гасторнисы были не плото-, а плодоядными. Получается, самый близкий аналог гасторнисов – дронт! Можно провести аналогию и с туканами или птицами-носорогами, но с поправкой на способности к полёту. Кроме прочего, это значит, что крупнейшими растительноядными существами Европы в палеоцене были птицы, то есть экосистема была во многом похожа на мадагаскарскую до прибытия туда людей. В общем-то, и географически ситуации схожи, ведь Европа была скорее архипелагом, чем материком. Кстати, до Северной Америки в палеоцене гигантские птицы ещё не добрались, так что там звери имели лучшие шансы на развитие своих вегетарианских наклонностей, которые они, собственно, успешно и реализовали. Учитывая ещё и архаичных страусов, тема больших нелетающих птиц была в палеоценовой Европе очень модной.
Если одни пернатые ограничивали наши возможности конкуренцией, другие просто нагло нас жрали! Конец палеоцена ознаменовался появлением сов – Berruornis orbisantiqui во Франции и Ogygoptynx wetmorei в Колорадо. Вот только наши предки избавились от гнёта злых динозавров, не прошло и десятка миллионов лет – как прилетела новая напасть! Думается, именно такие невзгоды были главными движителями нашей ранней эволюции: ночные хищники выдавили нас из ночного образа жизни, придали бодрости-прыгучести и сообразительности. Как знать, кабы не совушки – круглые головушки, может, так бы и до сих пор вместо нас-умничек по ветвям ползали приторможенные плезиадапиформы…
Конечно, главное достижение палеоцена – расцвет млекопитающих. Случился он не сразу, но очень быстро. Первый десяток миллионов лет кайнозоя звери были ещё очень мелкими, довольно однообразными и трудноразличимыми, но прогресс шёл небывалыми темпами. Вымирание коснулось млекопитающих как-то очень избирательно: кто-то исчез, кто-то сократился в числе и разнообразии, а кое-кто и преуспел.
Как ни странно, даже в палеоцене по углам планеты можно было найти остатних реликтов мезозоя и едва ли не палеозоя. Так, в Канаде до конца эпохи дожил Chronoperates paradoxus – симметродонт, примитивный настолько, что поначалу его приняли даже за распоследнего зверозубого-цинодонта! В принципе, современные однопроходные-яйцекладущие тоже ведь являются подобным осколком былого, даже более примитивным, чем симметродонты.
Кстати, палеоценовые однопроходные нам тоже известны, и – неожиданно! – из Аргентины, где жил родственный утконосу Monotrematum sudamericanum. Трудно судить о животном по трём молярам, но в этом случае важнее география: зоогеографически вырисовывается соединение Австралии, Антарктиды и Южной Америки в мелу и, возможно, палеогене.
В отличие от всегда редкостных однопроходных, аллотерии-многобугорчатые Multituberculata в палеоцене продолжали процветать. Вымирание их не просто не особо затронуло: в палеоцене число родов стало даже больше, чем в позднем мелу! С начала кайнозоя и до появления грызунов и нормальных приматов многобугорчатые оставались едва ли не главными растительноядными во многих фаунах. Их универсальная зубная система, отточенная до совершенства ещё с середины мезозоя, гарантировала успех даже с учётом общей примитивности группы. Долотовидные резцы для разгрызания, гребнеподобные премоляры для разрезания и плоские широкие моляры для пережёвывания вместе с мощными короткими и высокими челюстями – идеальная схема, которая неспроста изобреталась много раз параллельно в самых разных группах животных. Имея отличный задел, многобугорчатые эволюционировали в палеоцене едва ли не быстрее прочих млекопитающих. Например, ещё в самом начале палеоцена монтанский Taeniolabis taoensis имел пятнадцатисантиметровый череп и предположительно мог весить от двадцати до аж больше сотни килограмм! Прочие млекопитающие достигли таких показателей сильно позже. Едва ли не лучше прочих изучен Ptilodus: это было древесное существо с цепким хвостом для подвешивания и разворачивающейся назад стопой для лазания по стволам вниз головой. Очевидно, что конкуренция нашим предкам была весьма основательная; возможно, именно благодаря успеху многобугорчатых приматы с самого начала не скатились до специализированной растительноядности, не то эволюции нашего интеллекта мог быстро настать конец.

Ptilodus
Конечно, была от многобугорчатых и другая явная польза: они были не очень быстрыми и притом вкусными. В погадках хищных птиц из палеоценовых отложений Внутренней Монголии найдены кости и шерсть многобугорчатых Lambdopsalis bulla. Стало быть, хищным существам было кого ловить, тогда как более шустрые приматы могли оставаться в относительном покое (правда, именно в этой фауне приматов вовсе не было, но ведь хищники ловили многобугорчатых и в Северной Америке).
Многобугорчатые жили и в Южной Америке, откуда известна их специфическая форма Sudamerica ameghinoi, иногда выделяемая в группу Gondwanatheria. Эти звери развили высокие коронки, приспособленные для жевания травы. В Южной Америке уже были саванны, и многобугорчатые выходили в них? Пока это под вопросом.
Вообще, Южная Америка была заповедником всяких неведомых зверей. Удивительный реликт юрских и меловых времён – аргентинский Peligrotherium tropicalis из группы Dryolestida. Реликт реликтом, а вырос он до размеров собаки, став самым большим представителем дриолестид, причём перешедшим на растительноядность. Потомки его продержались на материке ещё довольно долго.
Сумчатые пережили позднемеловую катастрофу не идеально: огромное число линий прервалось, от былого разнообразия остались ошмётки. Вероятно, это дало отличный шанс плацентарным, которые, при всей своей прогрессивности, в мезозое не особо-то преобладали. Но и для сумчатых нашлось своё уютное местечко – целая Южная Америка, где в отсутствие плацентарных сумчатые развернулись в полную силу – уже в палеоцене они представлены тут несколькими отрядами. Правда, на взгляд обычного человека все южноамериканские опоссумоподобные зверушки выглядят до уныния однообразно и на фоне впечатляющих черепах, змей и крокодилов совершенно теряются, но и среди них можно найти оригиналов.
Самыми перспективными были, вероятно, боливийские Pucadelphys andinus, жившие в самом начале палеоцена. В местонахождении Тиупампа на пятачке менее чем в квадратный метр обнаружены останки 12 опоссумов, а в трёх метрах на таком же маленьком участке – ещё 23 особей, лежащих плотной кучкой! Очевидно, с ними произошло что-то нехорошее (скорее всего, они тихо-мирно сидели в норках на берегу озера, никого не трогали и погибли во время быстрого наводнения), но важно само скопление: очевидно, пукадельфисы были как минимум очень терпимы к присутствию рядом других особей, а как максимум – сколько-нибудь социальны. Косвенно о том же свидетельствует половой диморфизм: самцы пукадельфисов были куда более мордастыми и клыкастыми, чем самки. Обычно такое происходит из-за повышенной конкуренции между самцами за самок или даже гаремности. Из всей толпы определить пол удалось у двенадцати самок и шести самцов: соотношение «один к двум» может тоже говорить о повышенной социальности; как минимум четыре подростка показывают, что в группах уживались особи разного возраста. Судя по находке скопления из шести скелетов других опоссумов Andinodelphys cochabambensis в том же местонахождении, подобный образ жизни не был таким уж исключением. Здорово, что для синхронных приматоморфов и даже более поздних приматов у нас нет никаких доказательств хоть какой-нибудь общественности. Выходит, сумчатые в начале палеоцена даже слегка обогнали наших предков по уровню интеллекта! Если бы не недостатки системы размножения, глядишь, сейчас потомки южноамериканских опоссумов читали бы о вымерших пургаториусах и изучали реликтовых приматов – «живых ископаемых» дремучих палеогеновых времён.
Некоторые опоссумоподобные твари – Allqokirus australis и Mayulestes ferox – в палеоцене ещё не особо выделялись из общей серой массы (хотя в фауне Тиупампы они уже были самыми крупными сумчатыми), но начали претендовать на большее. Ближайшее будущее их было великолепно, ведь они стояли в основании группы Sparassodonta, или Borhyaenoidea, – главных хищников Южной Америки до появления кошек. В окружении крокодилов и при отсутствии плацентарных хищников плотоядные получаются и из сумчатых. Начав с аналога куницы – древесной, с очень подвижными лапками, да ещё и цепкохвостой, спрассодонты в последующие времена достигли размеров и облика тигра!
Среди прочих невзрачных палеоценовых южноамериканских сумчатых зверьков на первый взгляд ничем не выделяется Khasia cordillerensis. На самом же деле, эта скучная «мышка» весьма важна для зоогеографии, так как относится к отряду Microbiotheria. Их потомки – соневидные опоссумы Dromiciops gliroides – и сейчас живут в горных лесах юго-запада Южной Америки. Это единственные южноамериканские сумчатые, близкородственные австралийским сумчатым, отчего даже объединяются с ними в гиперотряд Australidelphia. Так, через древних и современных тварей прослеживаются связи материков.
Плацентарные начала палеоцена многочисленны, но почти всегда фрагментарны и трудноразличимы. Среди самых примитивных – Cimolesta – выделяется универсальностью Cimolestes: мало того, что он без проблем перешёл из мела в палеоцен, так ещё и найден и в Северной Америке, и в Боливии, и в Марокко! Некоторые авторы склонны разделить цимолестов на несколько родов, но разница между ними получается уж очень формальной. Цимолесты настолько усреднённо-примитивны, что просто наверняка являются предками прочих отрядов плацентарных. Правда, в палеоцене потомки уже существовали, так что цимолесты кайнозоя были «живыми ископаемыми». Из современных зверей на цимолестов максимально похожи опоссумы, ласки и землеройки, неспроста, например, цимолест Palaeoryctes считался древнейшим насекомоядным.
Из подобных генерализованных тварей в ускоренном темпе стали возникать самые разные варианты: насекомоядные, апатемииды, пантодонты, тиллодонты, тениодонты, кондиляртры, чуть позже – пантолесты, анагалиды, грызуны, копытные и хищные. Всё это изобилие можно условно разделить на несколько основных стилей жизни: мелкие насекомоядные зверьки, сравнительно крупные наземные растительноядные, мелкие растительноядные и хищники. Каждое из этих направлений реализовалось в нескольких альтернативных версиях.
Более специализированные, чем цимолесты, но тоже очень примитивные зверушки – Leptictida также успешно миновали границу мезозоя и кайнозоя. Состав их разными палеонтологами видится сильно по-разному. Например, монгольский Praolestes рассматривается либо как лептиктид, либо как примитивное насекомоядное.
Насекомоядные неплохо представлены в палеоцене: уже в начале эпохи в Китае появляются землеройкоподобные Carnilestes palaeoasiaticus, C. major и Prosarcodon lonanensis, а позже подобных зверушек становится ещё больше. План строения и образ жизни землероек с тех пор, конечно, тоже поменялись, но на фоне приматов они выглядят так архаично, что на их примере можно наглядно представить облик пушистиков начала кайнозоя. Североамериканские позднепалеоценовые Litolestes ignotus, Leipsanolestes siegfriedti, Cedrocherus aceratus и C. ryani, возможно, уже были первыми ежами.
«Насекомоядные» – не самый удачный термин, так как используется для обозначения как типа питания, так и систематической группы, причём её рамки разными исследователями видятся по-разному. Былой отряд Insectivora ныне не рассматривается как единый: прыгунчики, тенреки и златокроты в современных системах обычно отделяются в Afroinsectivora, а щелезубы, кроты, землеройки и ежи составляют Lipotyphla. С ископаемыми зверьками всё гораздо запутаннее, так как древнейшие представители слишком похожи друг на друга, хотя явно в итоге дали очень различных потомков.
Мелкие насекомоядные зациклились на беспозвоночных – это и плюс, и минус. Хорошая сторона в том, что насекомых и червей обычно мало не бывает, так что даже зимой под снегом землеройки умудряются находить достаточно еды, чтобы не замерзать. Плохая сторона в том, что при мизерных размерах тело моментально остывает, так что даже в тропиках питаться приходится постоянно: как известно, землеройки в день съедают больше, чем весят сами, а если зверушка поголодает пару часиков, то почти гарантированно умрёт. В таком режиме существовать можно, но достичь чего-то большего, нежели ловля червяков и жуков, сложновато. Благо, план строения насекомоядных весьма прост, а стало быть – годен для прогресса и производства более крупных и сложных форм. В принципе, из чего-то очень похожего, только в древолазательной версии, произошли и мы.
Apatemyidae – очень своеобразная североамериканско-европейская группа, иногда выделяемая в отряд Apatotheria, а иногда присоединяемая к другим, в том числе цимолестам или плезиадаписовым приматоморфам, по ряду же признаков чрезвычайно похожая вообще на летучих мышей. Уже палеоценовые апатемииды – например Apatemys kayi, Jepsenella praepropera и Labidolemur soricoides – имели очень специализированное строение: первые нижние резцы стали огромными лежачими, направленными строго вперёд скобелями, а верхние – резко утолщенными вертикальными крючками; боковые резцы и нижние клыки вообще исчезли; передние премоляры превратились в огромные режущие гребни, прямо как у многобугорчатых, а моляры уменьшились. Такой зубной аппарат был заточен под выгрызание насекомых из трухлявых пней, как это делают современные полосатые кускусы Dactylopsila на Новой Гвинее и руконожки Daubentonia madagascariensis на Мадагаскаре (впрочем, бывалых морфологов смущает немалое сходство зубного аппарата лабидолемура с таковым землероек – пинцет из нижнего «бивня» и верхнего «крюка»: землеройки пни не грызут!). В последующем, с появлением дятлов, оборудованных куда более эффективными клювами, апатемииды оказались не у дел и вымерли, а кускусы и руконожки спаслись только исключительно благодаря тому, что коварные дятлы пока ещё не добрались до их отдалённых островов.
В конце эпохи возникла и совсем иная версия цимолестов – Pantolesta, причём сразу в нескольких версиях. Внешне ничем не примечательные Bisonalveus browni и B. holtzmani таки примечательны продольными бороздками на клыках, которые могут быть свидетельством развития ядовитых желёз (такая же бороздка найдена ещё на одном клыке неописанного палеоценового зверька из Альберты). Ядовитость возникала у млекопитающих считаное число раз. Бизоналвеусы были мелкими насекомоядными тварюшками, которым могло быть важно быстро убивать каких-то особо прытких или опасных насекомых; возможно же, бизоналвеусы ядом защищались от врагов. Правда, далеко не каждая бороздка на клыке гарантирует ядовитость. Специализированный вариант пантолестов – выдроподобные Palaeosinopa simpsoni из Северной Америки – успешно перешли в эоцен, на который и пришёлся расцвет этой группы.
Ближайшие родственники и, видимо, потомки цимолестов – Pantodonta. В палеоцене и начале эоцена это были главные растительноядные звери. На их примере здорово видна бурная эволюция при освобождении экологических ниш после вымирания динозавров. Правда, до сих пор загадкой остаётся география пантодонтов: примитивнейший и древнейший раннепалеоценовый представитель найден – неожиданно! – в Южной Америке, в Боливии: Alcidedorbignya inopinata весила полкило и имела довольно-таки генерализованные зубы, что не исключает некоторых специфических специализированных черт. Чуть более поздние пантодонты жили в Северной Америке и Азии, где прослеживается чудесный ряд от Archaeolambda (0,7–1 м, 7–15 кг) через Pantolambda (1,2 м, 45 кг) к Barylambda (2,5 м, 600–650 кг), Coryphodon (2,5 м, 500 кг) и Titanoides (2,2–3 м, 90–200 кг).
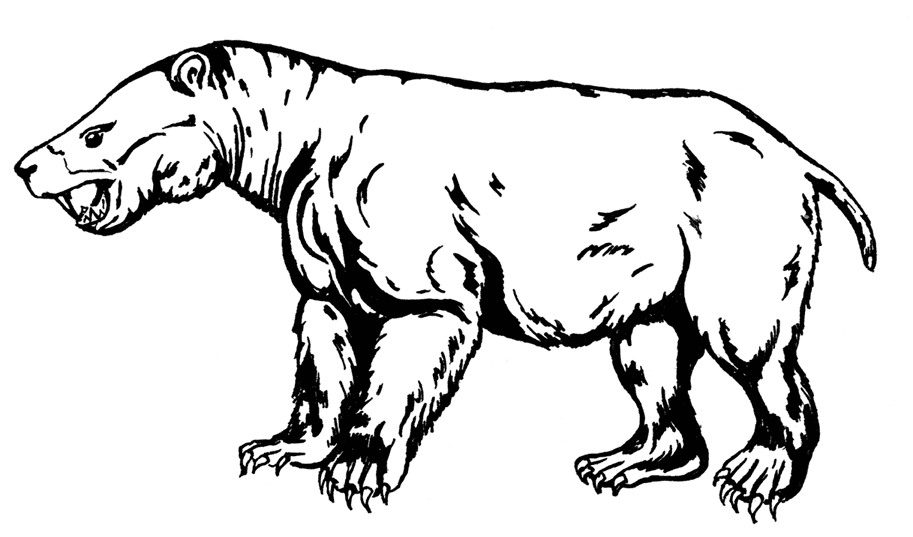
Titanoides
Кроме размеров, менялся и внешний облик: археолямбда была похожа скорее на кошку и ещё могла быть всеядно-насекомоядной, пантолямбда напоминала мордастого барсука, барилямбда с её махонькой головёшкой, высокими ногами и толстым, почти кенгурячьим хвостом – гигантского наземного ленивца, более продвинутый корифодон – тапира и даже более – карликового бегемота, а титаноидес – саблезубую помесь бегемота с медведем. У всех них были мощные передние и задние ноги с пятью растопыренными пальцами с округлыми ногтеподобными копытцами, причём от ранних форм к поздним происходил переход от стопо- к пальцехождению. Примитивными чертами было сохранение ключицы и разделённых костей предплечья и голени – наследие древесных предков (ранние виды, вероятно, ещё вполне успешно лазали по деревьям), хотя наземность крупных видов несомненна. Своеобразие пантодонтам придавали тупорылые морды с тяжёлыми челюстями и острыми гребнистыми зубами. Вот в зубах-то и была проблема: продольные гребни верхних входили в бороздки на нижних, так что боковые жевательные движения были практически невозможны, отчего, кстати, и скуловые дуги были довольно узкими. Пантодонты могли здорово стричь растения, но не умели нормально жевать, что в условиях климатического оптимума было не так уж важно, но при сокращении растительных богатств и появлении крутых конкурентов уже в конце эоцена стало фатальным. Другой сложностью были мозги: у пятисоткилограммового корифодона они весили несчастных 90 г. Для сравнения, у лошади того же размера мозг весит с полкило и немножко больше; да даже у овцы его масса – 140 г! Конечно, ранние копытные тоже вряд ли блистали интеллектом, но ведь и корифодон – уже далеко не самый ранний пантодонт!
В некоторых отношениях похожи на пантодонтов были Tillodontia, видимо, в силу родства, хотя по иным чертам они существенно отличались. У них тоже были мощные стопоходящие лапы, но пальцы заканчивались когтями; ключица и несросшиеся кости предплечья и голени по-прежнему отражают архаику. Главное отличие от пантодонтов – челюсти и зубы: боковые жевательные движения преобладали над продольными, отчего скуловые дуги широко расходились; резцы были резко увеличены, а вторые резцы были покрыты эмалью только спереди, не имели корней и постоянно росли, как у грызунов; уменьшенные клыки, как и резцы, имели долотовидную форму, а между ними и премолярами лежала широкая диастема. Такая зубная система имеет очевидную аналогию с грызунячьей, с поправкой на размер: тиллодонты, как и все в палеоцене, начав с маленьких зверушек, быстро доросли до вполне приличных масштабов. Тиллодонты появились в Китае (например, Meiostylodon zaoshiensis и другие мелкие примитивные виды), но довольно быстро оказались и в Северной Америке (например, Azygonyx – размером уже с енота). По-прежнему, проблемой были крошечные мозги; особенно здорово это видно при взгляде на череп сверху.
Ещё более быстроэволюционировавшая североамериканская группа – Taeniodonta; они появились ещё в мезозое; между прочим, опоссумоподобная Schowalteria clemensi была тогда в числе крупнейших млекопитающих. В палеоцене тениодонты начали с роющих всеядных Onychodectes tisonensis – зверя, похожего на крысу с приплюснутой головой и достаточно банальным набором зубов, – и довольно быстро подросли до Wortmania otariidens, у которой морда укоротилась, а нижняя челюсть стала очень тяжёлой, и далее – до Psittacotherium multifragum метровой длины, в полсотни килограмм весом и с совсем уж страшной нижней челюстью. Как и у тиллодонтов, тениодонты изобрели постоянно растущие передние зубы с эмалью лишь на передней стороне, с отличием в том, что главными грызущими зубами стали клыки, а в меньшей степени – боковые резцы. Огромными когтями на мощнейших лапах (опять пятипалых, опять с несросшимися костями предплечья и голени) тениодонты, видимо, выкапывали какие-то корни и клубни.
Таким образом, среди азиатских и североамериканских ранних растительноядных рисуется некоторое разделение труда: пантодонты стригли листья вертикальными движениями челюстей, тиллодонты грызли семена и жевали их поперечно, а тениодонты выкапывали корни, причём специализация во всех трёх группах нарастала скорейшими темпами.
Зубы млекопитающих отлично отражают их питание и образ жизни, недаром сказано: «по зубам их узнаете их». Что здорово, именно зубы-то обычно и сохраняются, так что по ним можно понять не только родство зверей, но и экологическую обстановку. Одонтология – великая сложная наука с массой нюансов; специалисты учатся этой премудрости всю жизнь. Однако некоторые базовые понятия вполне возможно изложить в двух словах. Особенно показательна форма моляров, ведь основная нагрузка приходится обычно именно на них.
Трибосфения – тип зубов с тремя острыми вершинками, расположенными по вершинам треугольника и соединенными гребнями, смыкающимися в замок с преобладанием давящих и ограничением жевательных движений, исходное примитивное состояние; в более продвинутой версии к треугольнику сзади добавляется пятка – талонид на нижних зубах и талон на верхних – для лучшего перетирания (у юрских Shuotheridia пятка добавилась спереди, что оказалось неудачной конструкцией и привело их к вымиранию); типична для опоссумов и землероек.
Секодонтия, или туберкуло-секториальность, – развитие трибосфении с добавлением новых острых вершинок, соединенных острыми гребнями и превращающих зуб в удлинённую режущую пилу; в крайнем случае формируется огромный хищнический зуб, берущий на себя главную нагрузку (он может образовываться и из премоляров, и из моляров; у креодонтов их было даже два); типична для хищных.
Бунодонтия – тип зубов с низкими округлыми бурками для раздавливания мягкой пищи; типична для свиней, похожа форма зубов у ежей. Возникали варианты, в том числе бунолесодонтия (бугорчато-лунчатые зубы), с превращением наружных бугров на верхних, а также внутренних бугров на нижних зубах в полулунные гребни для питания листьями деревьев у антракотериев, зиголофодонтия (она же циголофодонтия) с объединением бугорков в гребни у мастодонов.
Лофодонтия – тип зубов со слившимися в поперечные гребни бугорками для разрезания растительности; в процессе эволюции число гребней обычно растёт; типична для носорогов, лошадей и белок. Возникало много вариантов: от билофодонтии с двумя гребнями у тапиров, сирен, зайцеобразных и мартышек через трилофондонтию, тетралофодонтию и пенталофодонтию до полилофодонтии с множеством пластин у слонов и капибар.
Селенодонтия (лунчатые зубы) – тип зубов с потерей эмали на вершинах бугорков, отчего режущие эмалевые кромки превращаются в полумесяцы, а жевательные движения оказываются поперечными; типична для быков и оленей.
Брахиодонтия – низкокоронковость, исходное примитивное состояние; типична для свиней и белок.
Гипсодонтия – высококоронковость, специализированное состояние, приспособленное для питания большим количеством жёстких растений; типична для парно- и непарнокопытных и большинства грызунов.
Гипселодонтия – крайняя форма гипсодонтии с потерей корней и неограниченным ростом для перетирания большого количества малопитательной травы; типична для резцов и щёчных зубов полёвок и зайцев, клыков свиней, жевательных зубов эласмотериев.
Самой разнородной группой всеядно-растительноядных палеоценовых зверей были Condylarthra. Споры о их сущности не утихают с момента их открытия. Как только не группировали отдельные группы кондиляртр, какие только схемы родства их с иными млекопитающими не предлагали! И ведь, с одной стороны, часть представителей очень уж хорошо различима, но с другой – всегда находятся какие-то невнятные промежуточные формы, стирающие границы. В итоге единство группы находится под основательным сомнением, но и совсем отказаться от использования понятия «кондиляртры» никак не получается. Базовой версией и даже, по мнению некоторых палеонтологов, исходником для всех истинных плацентарных или как минимум копытных, можно считать Protungulatum. Этот североамериканский зверь успешно перешёл мел-палеогеновую границу и дал в палеоцене чудесный веер форм.
Французские Pleuraspidotherium aumonieri и Orthaspidotherium edwardsi с их длинными низкими плоскими головами, небольшими, но острыми клыками, клыкоподобными первыми премолярами и молярами, близкими к молярам копытных, были похожи скорее на пристукнутых сковородкой всеядных виверр. Возможно, их подобие предкам копытных конвергентно, а на самом деле они представляют самостоятельную тупиковую линию с мозаикой примитивных и продвинутых черт.
Сборная группа Hyopsodontidae включает мелких, преимущественно насекомоядных и хорошо лазающих по деревьям зверьков, например Paschatherium и Aletodon. Иные – клыкастые Loxolophus и Chriacus, Ectoconus и Periptychus – экологически были похожи на енотов и вполне могли составлять конкуренцию ранним приматам.

Chriacus
Предками непарнокопытных могли быть представители Phenacodontidae, например Tetraclaenodon и Ectocion, похожие на коротконогих плоскоголовых собачек-вегетарианцев.
Из кондиляртр формировались и необычные слонопотамы: в Китае и Монголии несколько видов Prodinoceras, а в Северной Америке Probathyopsis (в эоцене этот род дошёл и до Китая) представляют первые образцы группы Dinocerata (вообще, как обычно, их корни теряются в изобилии палеоценовых зверей и могут быть связаны как с разными кондиляртрами, в частности с арктоционидами, так и с пантодонтами, анагалидами, ксенунгулятами и много кем ещё). В будущем они станут весьма внушительными аналогами носорогов, а ныне были приземистыми коротконогими тварями невзрачной наружности, хотя могли похвастаться огромными клыками и уже доросли до трёх метров длины. Передние лапы у них были очень мощными пальцеходящими, неплохо приспособленными для копания, а задние – стопоходящими. Удивительно внешнее сходство ранних диноцерат и дицинодонтов – зверообразных рептилий перми и триаса. В отличие от нормальных растительноядных, ранние диноцераты практически не могли жевать ни в передне-заднем, ни в поперечном направлениях, так как верхние и нижние зубы смыкались в плотный замок. При этом жевательная мускулатура была крайне мощной. Получается, диноцераты могли очень сильно кусать, но только по вертикали. Судя по всему, они ещё не были полностью растительноядными и вместе с травой и корнями выкапывали из земли всяческих червей и слизняков.
Каким-то образом кондиляртроподобные твари проникли и в Южную Америку, где в Боливии, Бразилии и Аргентине известно немалое число их видов, причём довольно разнообразных. Из них особенно замечательна Lamegoia conodonta: её зубы внешне удивительно похожи на зубы высших приматов. По крайней мере некоторые из южноамериканских кондиляртр стали предками своеобразных «южноамериканских копытных», развернувшихся в полную силу уже в следующие эпохи.
Впрочем, некоторые уже успели доэволюционировать до более продвинутого состояния: примитивнейшими представителями Notoungulata стали аргентинские Simpsonotus major и S. praecursor – растительноядные зверушки, похожие на собак размером с кошку, чьи третьи верхние резцы превратились в подобие клыков; в группе Litopterna первой отметилась аргентинская Requisia vidmari, а родственный Notonychops powelli иногда выделяется в собственный отряд Notopterna; толстомордый Eoastrapostylops riolorense был первым среди Astrapotheria. Расцвет всех этих групп был ещё впереди.
В ранних южноамериканских кондиляртрах, нотоунгулятах и прочих зверях разобраться весьма сложно: находок-то много, но большинство из них – изолированные зубы, причём между крайними формами имеется масса промежуточных, так что провести границы не только видов, но даже более высоких систематических категорий весьма непросто. Например, среди прочего изобилия аргентинский палеонтолог Флорентино Амегино описал 3 отряда, 4 семейства, 8 родов и 16 видов, которые впоследствии оказались останками одного вида Henricosbornia lophodonta! И ведь невозможно упрекнуть Ф. Амегино в непрофессионализме – для южноамериканской палеонтологии он – альфа и омега. Просто задача действительно очень непростая.
Особняком среди южноамериканских странных животных стоят несколько видов Carodnia, выделяемых в самостоятельный отряд Xenungulata. Внешне и по размеру они были похожи на помесь капибары с тапиром: приземистые, с копытцами на пяти растопыренных пальцах, короткомордые, с колышкообразными резцами, большими клыками и молярами с поперечными гребнями. В своё время это были самые крупные южноамериканские растительноядные звери.
Впрочем, как бы ни были увлекательны южноамериканские звери, к нашей эволюции они имеют весьма косвенное отношение. Гораздо важнее для появления нас были изменения экосистем Африки, ведь именно там в последующее время было суждено сформироваться людям. К сожалению, про Африку-то мы знаем меньше всего: на большей части континента палеоценовых фаун пока вообще не найдено. Но есть одно местечко в Марокко – Улед Абдун, где найдено пусть и не очень много окаменелостей, зато очень показательных. Звездой марокканского палеоцена, без сомнения, является Ocepeia daouiensis: от неё сохранились два черепа, а также обломки челюстей и изолированные зубы. Этот зверёк размером с кошку – около 3,5 кг весом – выглядел по палеоценовой традиции весьма невзрачно и первоначально был зачислен в кашу кондиляртр, но суть, как обычно, в деталях. Оцепейя с её увеличенными пупырчато-гребнистыми молярами явно была листоядной. Её череп укорочен и массивен, с необычно и непонятно зачем развитыми воздушными полостями и широкой носовой полостью. Все эти признаки сыграли свои роли, когда потомки оцепейи стали прыгунчиками, тенреками, даманами, трубкозубами и златокротами, ведь этот зверёк представляет идеальную переходную стадию между насекомоядными и группой Afrotheria; одновременно при желании можно попытаться найти в оцепейе черты слонов и сирен. Любопытно, что по форме черепа и некоторым особенностям зубов, например уплощённой форме резцов и толстым клыкам, оцепейя конвергентно похожа на примитивных приматов. Получается, мы видим наглядный пример, куда могло занести нас – в аналоги трубкозубов и слонов! С другой стороны, выходит, что и даманы с тенреками имели шанс! К сожалению, пока мы не знаем, как выглядело тело оцепейи, но, судя по потомкам, вряд ли она умела хорошо лазать по деревьям. Вероятно, эта приземлённость и не позволила афротериям развить хватательную конечность, освоить орудия труда и стать разумными.
Как ни мало найдено в Марокко, но и тут можно проследить эволюцию: чуть более поздняя Ocepeia grandis в полтора раза крупнее по длине и втрое тяжелее.
Существует много вариантов подразделения плацентарных млекопитающих на группы. Судя по всему, конвергенция – независимое возникновение сходства в силу одинакового образа жизни – была самым обычным делом, так что старые схемы, ориентировавшиеся на простые внешние признаки, не отражают родства. Например, в группу Pachydermata – «толстокожие» – включались все звери с копытами, простым желудком и более-менее массивным сложением – слоны, носороги, бегемоты и лошади. Развитие морфологии и палеонтологии позволило сделать схемы более реалистичными. Намного большее значение оказалось за особенностями строения зубов и слуховой капсулы, а также расположения и формы отверстий для нервов и сосудов на основании черепа, поскольку они достаточно специфичны и очень разнообразны. Но на надотрядном уровне и такие ухищрения помогают мало, ведь большинство отрядов разошлись очень давно, едва ли не в мезозое, и былое родство сильно затёрлось миллионами лет специализаций, а предковые промежуточные формы бывают слишком невнятны или чересчур фрагментарны, а то и вовсе не найдены. Новое дыхание в систематику привнесли молекулярная биология и генетика. Тут, правда, есть свои сложности: во-первых, по разным биохимическим системам и генам схемы получаются порой сильно разными, во-вторых, конвергенция вполне бывает и в мире белков и генов, а в-третьих, у нас нет белков и генов ископаемых животных, кроме разве что самых недавних, плейстоценовых, да и то в виде редкостного исключения.
Но учёные не сдаются. Наработок и подходов в разных областях уже столько, что общая схема вполне вырисовывается.
По одной из самых распространённых версий плацентарные млекопитающие делятся на две-три большие группы. «Южная» – Atlantogenata, целостность которой под вопросом. Возможно, особняком ото всех плацентарных стоят неполнозубые Xenarthra. Гораздо более консолидированная подгруппа атлантогенат – Afrotheria, включающая Afroinsectiphilia (тенреки, златокроты, прыгунчики и трубкозубы), а также Paenungulata (даманы и хоботные с сиренами). «Северная» группа – Boreoeutheria из двух подгрупп. Первая – Euarchontoglires с Archonta (или Euarchonta; тупайи, шерстокрылы и приматы) и Glires (грызуны и зайцеобразные). Вторая – Laurasiatheria с более заковыристым составом: Lipotyphla (насекомоядные – щелезуб, кроты, ежи и землеройки) и Scrotifera. Последние имеют довольно спорное подразделение: Euungulata (непарнокопытные, мозоленогие, парнокопытные и китообразные) и Ferae (панголины и хищные с ластоногими), а рукокрылые зависают где-то между ними.
Получается, с самых ранних времён плацентарные разделились пополам, и в каждой половине возникали во многом похожие варианты, эволюционные дубли. С учётом же сумчатых и вымерших южноамериканских зверей получилось аж четыре альтернативных версии млекопитающего мира: австралийский, южноамериканский, африканский и евразийско-североамериканский. Из них один – южноамериканский – почти полностью исчез, второй – австралийский – сохранился только благодаря изоляции, а третий – африканский – в виде добавки влился в самый успешный четвёртый. Степень успеха, как кажется, пропорциональна размеру территории и разнообразию условий.
Правда, далеко не все зоологи согласны с подобным делением млекопитающих. Многие указывают, что анатомических оснований объединения афротериев как-то вроде и нет, две их главные группы – мелкие насекомоядные Afroinsectiphilia и большие растительноядные Paenungulata – морфологически чётко разделены уже в конце палеоцена и начале эоцена. Ценность оцепейи в немалой степени в том и состоит, что она образует смычку между двумя половинами группы, выделенной первоначально только по генетическим данным. С другой стороны, оцепейя жила в одно время с уже готовыми хоботными, так что никак не может быть их прямым предком.
Другой ранний афротерий из тех же марокканских слоёв – Abdounodus hamdii. Как и оцепейя, изначально он был описан как кондиляртра, причём очень похожая на боливийских Mioclaenidae типа Pucanodus gagnieri, что любопытно географически. С другой стороны, оцепейя и абдунодус напоминают китайско-монгольских палеоценовых Phenacolophidae типа Minchenella grandisu, Phenacolophus fallax и нескольких других похожих зверушек, представляющих, возможно, первую версию загадочной группы афротериев Embrithopoda, в следующие эпохи выплывающей в Румынии, Турции и Северной Африке. А ведь поздние эмбритоподы были отдалённо похожи на слонов, которые вроде как родственники оцепейи и абдунодуса… Но вроде как афротерии – группа африканская, при чём же тут Китай и Монголия? В целом ясно, что палеоценовая масса кондиляртроподобных тварей ещё будет неоднократно пересматриваться, слишком пока всё это запутано.
В том же Марокко и в то же время жил и потенциальный предок тенреков и златокротов Todralestes variablis: его острые зубки явно больше нацелены на насекомых, чем растения. Как и у многих палеоценовых зверьков, признаки тодралестеса настолько обобщённые, что его первоначально отнесли к отряду Proteutheria, в который иногда зачисляют всех ранних невнятных насекомоядных, начиная с юрамайи через цимолестов до пантолестов и апатемиид.
Тут же в Марокко, даже в одном местонахождении с оцепейей и абдунодусом, найдены уже и первые готовые хоботные. В палеоцене есть уже два их последовательных варианта: Eritherium azzouzorum и, в более высоком слое с плавным переходом в эоцен, – Phosphatherium escuillieri. Начало самых гигантских наземных животных современности было скромным – похожие на тапиров зверьки размером с карликовую свинку. В отличие от потомков, зубная формула эритерия самая что ни на есть генерализованная, без уменьшения числа зубов, но со слабым намёком на будущие изменения в слоновью сторону; бунодонтные зубы говорят о питании мягкой растительной пищей, а огромная носовая полость может быть приспособлением к жизни в воде. От фосфатерия тоже сохранилось не так уж мало, так что мы знаем о его длинной и широкой носовой полости, намекающей на будущую хоботастость, увеличении резцов, намекающей на будущие бивни, переходе к лофодонтии – намёке на листоядность, а также об утере одного нижнего резца и одного премоляра по сравнению с эритерием. Фосфатерий подрос как минимум в полтора раза до длины аж 60 см и веса 10–15 кг – маловато для слона, но в самый раз для великого слоновьего предка.
Коли уж развелось столько сочных зверьков-вкусняшек, не могли не появиться хищные звери. Крокодилы и птицы, конечно, тоже не голодали, но исходная насекомоядность млекопитающих прямо-таки напрашивалась на создание крупных плотоядных. Систематические взаимоотношения ранних хищников, как обычно, крайне запутаны, клыкастые цимолесты, кондиляртры, мезонихии, гиенодонты, креодонты и хищные комбинируются разными авторами в самых причудливых комбинациях.
Существует мощная разница между понятиями «хищники» и «хищные». «Хищники» – это образ жизни и тип питания, таковыми могут быть хоть раки, хоть звери, хоть бы и растения. «Хищные» же – Carnivora – это вполне конкретный отряд млекопитающих, причём некоторые его представители по типу питания вполне себе вегетарианцы, все ведь знают о пандах.
Всеядные кондиляртры Deltatherium и Arctocyon размером с волка, с их вытянутой хищной мордой с большими клыками, выглядели серьёзно и, вероятно, действительно родственны хищным и одновременно, как ни странно, парнокопытным. От тех и других их отличал очень примитивный мозг – маленький, с огромными обонятельными долями и, вероятно, совсем без извилин. Впрочем, для обмана тогдашних травоядных интеллекта недоволков-микроцефалов вполне хватало, да и, судя по молярам, растения были существенной частью их рациона. Совсем уж саблезубым стал Mentoclaenodon acrogenius – едва ли не самый опасный хищник палеоцена. Разные виды рода Arctocyon, что интересно, жили и в Европе (например, A. primaevus), и в Северной Америке (например, очень большой A. mumak). Иногда подобные всеядные кондиляртры, например североамериканский Escatepos campi, уже считаются первыми креодонтами.
Иные некрупные острозубые североамериканские Tytthaena lichna и T. parrisi, а также несколько чуть более продвинутые Dipsalodon matthewi, D. churchillorum и Oxyaena transiens тоже претендуют на звание древнейших креодонтов, а порой ставятся в основание самостоятельного отряда Oxyaenodonta, вроде бы родственного хищным и отличающегося, в числе прочего, потерей третьих моляров и разворотом второго верхнего моляра в поперечном направлении. Показательно, что титтэны найдены в одних слоях с плезиадаписами, которым ещё вряд ли могли навредить, но на детёнышей которых вполне могли поглядывать с гастрономическим интересом. Иные их ближайшие родственники – североамериканские Palaeonictis peloria – в самом конце эпохи уже вымахали до размеров маленького медведя, став самыми большими хищниками палеоцена. Судя по мощности нижней челюсти, очень широким мыщелкам нижней челюсти, а также сравнительно широким и притуплённым щёчным зубам, палеониктисы могли заниматься падальничеством, разгрызая кости.
Во Внутренней Монголии обнаруживается Prolimnocyon chowi – один из древнейших представителей группы Hyaenodonta (учитывая соединения материков, не странно, что в эоцене несколько видов того же рода жили в Северной Америке). Гиенодонты иногда включаются в креодонтов, с чем, впрочем, некоторые палеонтологи не согласны; в последнем случае они рассматриваются как самостоятельная группа.
Creodonta – группа примитивных хищников, главных плотоядных первой половины кайнозоя. Возникли креодонты, видимо, в Азии, почти сразу оказались в Северной Америке, а после добрались и до Европы, и до Африки. Впрочем, есть и другое мнение, согласно которому креодонты – африканская группа, родственная панголинам. Рамки группы рассматриваются разными палеонтологами сильно неодинаково. Иногда в креодонтов включались некоторые кондиляртры, мезонихии и отдельные семейства, которые теперь включаются в хищных. Чаще в первом приближении креодонты делятся на семейства Hyaenodontidae и Oxyaenidae, которым иногда придаётся ранг самостоятельных отрядов Hyaenodonta и Oxyaenodonta. Оксиенодонты чаще были мелкими вытянутыми лазающими хищниками, а гиенодонты – крупными собако- и гиеноподобными бегающими, хотя в обеих группах разнообразие форм очень велико.
В Африке шли параллельные процессы. Марокканские Lahimia selloumi и Tinerhodon disputatum представляют промежуточную фазу между насекомоядноподобными цимолестами и Hyaenodonta, а иногда прямо зачисляются в состав последних и при определённом систематическом раскладе считаются древнейшими креодонтами Африки, то есть представляют зарю хищников. У них ещё не было хищнических зубов, а третьи моляры никуда не делись. Это были очень мелкие зверьки, напоминавшие скорее ласку и внешне совсем не походившие на своих более поздних чудовищных родственников.
Одновременно, причём с самого начала палеоцена, в Азии, Северной Америке и Европе хищники формировались и на базе копытных: отряд Mesonychia родственен с одной стороны кондиляртрам, с другой – парнокопытным и китам, с третьей, вроде бы, креодонтам, но это не точно. Как обычно, близость палеоценовых зверей к общим генерализованным предкам заставляет сомневаться в последовательности филогенетических ветвлений. Ранние китайские Hukoutherium ambigum и H. shimemensis были уже не такими уж мелкими собакоподобными клыкастыми хищниками, бесчисленные виды Dissacus распространились по трём материкам и выросли до масштаба шакала, а североамериканский Ankalagon saurognathus – с медведя, причём у последнего вида некоторые особи вдвое меньше, что намекает на резкий половой диморфизм. При всей своей хищности, мезонихии имели на пальцах копыта, хотя бы и когтеподобные.
Кровожадные поползновения частично реализовались на основе сумчатых и кондиляртр, но уже в палеоцене возникли и представители отряда хищных Carnivora (между прочим, раньше всех более-менее явно плотоядных раннекайнозойских плацентарных запихивали в группу Miacoidea, но её рамки оказались слишком резиновыми, так что она оказалась типичной «мусорной корзиной»). Первыми оказались Viverravidae, например множество видов рода Protictis, Ictidopappus mustelinus и некоторые другие их родственники. Судя по исчезновению третьего моляра, специализация уже в палеоцене успела зайти довольно далеко. Название виверравид отражает реальность: они правда были похожи на виверр (с другой стороны, виверры, очевидно, сохранили очень примитивный план строения). Строго говоря, они питались всё ещё в основном насекомыми, но надо же было с чего-то начинать. Развитие подобных существ к границе эоцена стало мощнейшим фактором нашей эволюции: ловкие древолазающие хищники с мало-мальски рабочими мозгами начали очень эффективно отлавливать приматоморфов-плезиадаписовых. Медленно лазающие и наземные формы приматоморфов с когтями на пальцах и маленькими мозгами оказались очень уязвимы и быстро исчезли, а выжили только самые шустрые.
Одна из важнейших особенностей хищников – хищнические зубы. Они формировались несколько раз независимо на основе разных зубов. У хищных это четвёртый верхний премоляр и первый нижний моляр, у гиенодонтов второй верхний моляр и третий нижний моляр, у оксиенодонтов первый верхний моляр и второй нижний моляр, у креодонтов первый и второй верхние моляры и второй и третий нижние моляры.
Своим путём пошли некоторые сумчатые хищники: у австралийского Thylacoleo хищнические зубы в виде гигантских лезвий сформировались из третьих верхнего и нижнего премоляров и, в меньшей степени, первого нижнего моляра, а роль клыков взяли на себя первые резцы.
Иные родственники пошли своим путём. Escavadodon zygus из раннего палеоцена Северной Америки представляет первую стадию странной группы Palaeanodonta. Судя по зубам, палеанодонты развились из лептиктид и в чуть меньшей степени родственны пантолестам, а из современных животных максимально близки панголинам и хищным. Эскавадодон был приземистым существом полуметровой длины (причём половину её занимал хвост) весом от полукилограмма до килограмма, с довольно мощными – толстыми и мускулистыми – лапами и сравнительно мелкими зубами. Вероятно, это были специалисты по выкапыванию червей и насекомых из почвы. Зубы ранних палеанодонтов сочетают противоречивые черты: у эскавадодона и позднепалеоценового китайско-монгольского Ernanodon antelios на тонкой, узкой и низкой нижней челюсти ещё сохранялись резцы, клык, четыре премоляра и три совсем махоньких моляра, то есть зубная формула была полной, но размеры зубов резко уменьшились. Если у эскавадодона зубы выглядели ещё более-менее привычно, у эрнанодона почти все зубёшки, кроме второго моляра, имели лишь один корень и далеко отстояли друг от друга в виде кривеньких колышков; зато клыки были длинные. Тупорылой мордой эрнанодон был похож, видимо, на ленивца.
Исходно в челюсти млекопитающих было пять премоляров. В дальнейшем их число уменьшалось, но в разных линиях по-разному. У лептиктид и палеанодонтов первым исчез третий премоляр, а у большинства прочих, включая приматов, – сначала пятый, потом первый, а у продвинутых – и второй.
Линия мелких всеядно-растительноядных зверей реализовалась в нескольких версиях. В Азии появились сразу две конкурирующие группы – анагалиды и грызуны. С самого начала они разошлись по приспособлениям зубов, хотя изначально вроде бы достаточно родственны. Anagalida сохранили примитивный набор зубов и острые бугорки, но коронки стали высокими, а премоляры – моляроподобными. Вероятно, анагалиды выкапывали еду из земли: оттого для увеличения подвижности в их предплечьях и голенях кости оставались несросшимися, пальцы на лапках были снабжены копытцеобразными коготками, а стёртость зубов явно повышена. Судя по высокой восходящей ветви нижней челюсти, анагалиды достаточно старательно жевали свою пищу, какой бы она ни была. Анагалиды появились в палеоцене Монголии и Китая (например, Stenanagale xiangensis и Pseudictops lophiodon) и сохранялись тут до олигоцена. В ранних местонахождениях они численно преобладают над многобугорчатыми и первыми грызунами.
Несколько позже на первый план вышли грызуны Rodentia (или, в более широком понимании, – Glires). Самые первые их представители объединяются в Eurymylidae, а иногда даже выделяются в собственный отряд Mixodontia. Они узнаваемы по увеличенным резцам и зазору-диастеме между резцами и щёчными зубами. Миксодонты очень быстро набрали обороты и дали огромное количество форм: например, позднепалеоценовые китайские Sinomylus zhaii, Eomylus borealis, E. bayanulanensis, Palaeomylus lii, монгольские Eurymylus laticeps и Asiaparamys shevyrevae, а также другие их родичи. Показательно, что в Северном Китае остатки миксодонтов в основном обнаруживаются в погадках хищных птиц: с самого своего появления грызуны стали едва ли не главной едой многих пернатых. Любопытно, что в этих же фаунах нет плезиадаписов, приматов и летучих мышей. Хорошо, что у наших предков была возможность эволюционировать в других местах, и к моменту проникновения грызунов в Северную Америку и Европу тамошние приматы-аборигены уже достигли достаточного уровня, чтобы выдержать конкуренцию с понаехавшими.
Arctostylopida в лице Arctostylops steini в Северной Америке и множества азиатских видов (например, Sinostylops promissus в Китае и Kazachostylops occidentalis в Казахстане) – ещё одна загадочная группа, родственная то ли грызунам, то ли вообще южноамериканским нотоунгулятам. Внешне похожие на даманов, с укороченной скуластой мордочкой и неспециализированными зубами, они никогда не были многочисленными и, видимо, проиграли конкуренцию обычным грызунам. Последние арктостилопиды вымерли уже в первой половине эоцена.
Ещё более близкие наши родственники – шерстокрылы Dermoptera. Появились они в Северной Америке. Уже в начале эпохи известен Dracontolestes aphantus, в середине – Mixodectes malaris и M. pungens, а в конце – Planetetherium mirabile, Worlandia inusitata и Plagiomene zalmouti. В отличие от современных шерстокрылов, Mixodectidae и Plagiomenidae имели резко увеличенные долотовидные резцы, как у многобугорчатых, грызунов и некоторых плезиадаписовых. Умели ли палеоценовые шерстокрылы парить на летательных перепонках, как это делают современные, мы не знаем, с большой вероятностью – нет. Как и у других групп, корни шерстокрылов растворяются в ранних плацентарных, а первые представители определяются с трудом и сомнениями. Например, целое семейство Cyriacotheriidae – Cyriacotherium argyreum, Presbytherium rhodorugatus и P. taurus – одними палеонтологами причисляется к пантодонтам, а другими – к шерстокрылам.
А где же люди? Первые приматы
Существует универсальный принцип для прояснения любого вопроса: надо идти к истокам. Так и с нашим происхождением – все самые главные наши качества были заложены у самых первых предков. А среди приматоморфов первыми были плезиадаписовые Plesiadapiformes, а среди них – пургаториусы Purgatorius.

Purgatorius
Пургаториусы – поистине Великие Предки! Их крошечные челюстюшечки и зубёшечки найдены в отложениях начала палеогена Северной Америки. К сожалению, до сих пор непонятно, кто был их предками в конце мела. И, как ни странно, дело не в недостатке данных, а в избытке: среди великого изобилия позднемеловых млекопитающих есть много очень похожих линий, но их детальное родство пока выявляется с большим трудом – слишком они похожи друг на друга и одновременно отличаются от своих предков. Очевидно, на границе эр эволюция дала основательный рывок, который ещё предстоит описать. Не исключено к тому же, что предков наших предков стоит искать не в североамериканских фаунах, а в европейских или азиатских, а те на границе мела и палеогена изучены пока ещё недостаточно.
Кроме пургаториусов, есть и иные претенденты на звание Великого Предка – загадочные адаписорикулиды Adapisoriculidae. Этих существ относили и к приматам, и к сумчатым, и к насекомоядным, и, что закономерно, выделяли как группу «неясного родства». Их зубы сильно отличаются от зубов плезиадапиформ – широкой стилярной полкой, как у опоссумовых, и отсутствием гипокона, как у опоссумовых и шерстокрылов, тогда как плечевые и бедренные кости, напротив, схожи с плезиадапиформовыми, а кости предплюсны – и с плезиадапиформовыми, и с шерстокрыловыми. Как и большинство ранних млекопитающих, адаписорикулиды были насекомо-фруктоядными, хотя отличие их диеты от ближайших конкурентов пока неведомо.
Парадоксально, но адаписорикулиды известны по довольно полным материалам, превосходящим данные по пургаториусам, но просто хуже изучены.
Если адаписорикулиды не были нашими прямыми предками, то они представляют отличный пример конвергенции – сходства, возникающего в разных группах в силу одинакового образа жизни. В то время как в Северной Америке и Восточной Азии эволюционировали приматоморфы, в Индии, Северной Африке и Западной Европе – адаписорикулиды. В чём они проигрывали – непонятно. А может, это именно их потомки читают нынче эти строки, а пургаториусы – просто чересчур разрекламированные самозванцы?..
Зато пургаториусы известны достаточно хорошо. Ни одного целого скелета пургаториусов пока не найдено, но обилие челюстей, зубов, пяточных и таранных костей позволяет в первом приближении прикинуть их облик. Мы знаем, что это были малюсенькие зверушки – что-то около бурозубки или мыши размером, а внешним видом, вероятно, средние между землеройкой и тупайей. Судя по зубам, пургаториусы были всеядными, причём разные виды немножко отличались. Например, P. janisae и P. mckeeveri принципиально похожи друг на друга, но, как обычно, весь интерес в деталях. У первого зверька моляры сравнительно низкие и короткие, с острыми бугорками и гребнями, а у второго – выше и длиннее, но с более уплощёнными бугорками и более округлыми гребнями. Это говорит о том, что P. janisae был более насекомоядным, тогда как P. mckeeveri мог, кроме насекомых, лопать и фрукты, и что попало.
Ясно, что в фауне с пургаториусами есть и другие зверушки. И вот сравнение диеты разных позднемеловых и раннепалеоценовых млекопитающих рисует чудесную картину.
В конце мела, на излёте динозавровой эпохи главными североамериканскими фруктоядами были сумчатые Glasbius twitchelli. Не исключено, что эти недоопоссумы не давали развиться нашим предкам. Но метеорит упал не зря: экологические ниши подосвободились, и пургаториусы стали всеядными. Показательно, что уже некоторые даже самые ранние приматы стремительно уходили в пищевые специализации: вдвое более крупный Ursolestes perpetior стал совсем насекомоядным, тогда как Paromomys farrandi и Pandemonium dis полюбили фрукты. Purgatorius mckeeveri оказывается самым универсалом, занимая на палеодиетологических графиках практически строго среднее положение. Вместе с тем, иные тогдашние звери – сумчатые и предки современных копытных и хищных – были гораздо более выраженными насекомоядами.
Судя по костям ножек, только пургаториусы среди первых кайнозойских зверей наловчились здорово лазить по деревьям, что определило всю нашу дальнейшую судьбу.
Таранные и пяточные кости пургаториусов, как и положено костям Великих Предков, сочетают очень примитивный план строения с мелкими прогрессивными деталями, намекающими на грядущий триумф. Таранные кости укороченными и расширенными пропорциями максимально схожи с костями кондиляртры Protungulatum и отличаются от вытянутой формы шерстокрылов, тупай и полуобезьян. Вместе с тем форма и степень слияния суставных поверхностей таранной кости пургаториуса уже куда как продвинутая и как раз больше похожа на приматоморфную.
Аналогично на пяточной кости: у пургаториуса она очень широкая в передней части, как у кондиляртр, тогда как в продвинутой версии приматов суживается. А вот суставные поверхности пургаториусовой пятки скорее приматные. Увеличение суставных фасеток и слияние отдельных маленьких фасеток в общие большие отражает увеличение подвижности стопы. Судя по всему, возросли возможности поворота ножки, что особенно актуально при древолазании и прыгании по веточкам.
Забавно, что у нас общая форма таранной кости из-за спуска на землю и потери прыгательных способностей вторично вернулась к точно тем же пропорциям, что были присущи предкам копытных и пургаториусам десятки миллионов лет назад! Не смотрите на тупайю свысока: её таранная кость прогрессивнее, чем у Вас!
Всеядность вкупе с древесностью дали нам подвижность, цепкие ручки, преобладание зрения, безопасность жизни, K-стратегию размножения с минимумом детёнышей и максимумом выживания, обучаемость, сообразительность, социальность и в итоге стали залогом разумности!
Чрезвычайно показательно соотношение разных видов в последовательных фаунах. В конце мела известен всего один зуб пургаториуса, причём есть сомнения – не затесался ли он в этот слой из более поздних?
Через 25–80 тысяч лет после окончания мезозоя – начала кайнозоя никаких приматоподобных существ в Монтане совсем не видать, причём фауны-то довольно богатые. Либо пургаториусы просто ещё не возникли, либо позднемеловая катастрофа лишила местность всех деревьев, либо пургаториусы пришли сюда позже из других мест.
А вот спустя ещё каких-то несчастных двадцать-пятьдесят тысяч лет ситуация резко меняется.
На севере Монтаны, на склонах Кордильер, стояло одинокое ранчо… Впрочем, нет, это другая песня, у нас не про гусей, а про пургаториусов. Так вот, на севере Монтаны, в довольно-таки безрадостной местности, палеонтологи накопали некоторое количество окаменелостей. Отложения этой пустоши весьма любопытны тем, что сформировались 66 миллионов лет назад, в конце эпохи пуэркия, то есть самого начала кайнозоя, всего через 105–139 тысяч лет после окончания мезозоя, то бишь вымирания динозавров. И вот же радость – тут нашлись зубы как минимум двух видов древнейших приматов из группы плезиадапиформ – Purgatorius janisae и P. mckeeveri.
250–328 тысяч лет опосля падения метеорита плезиадапиформы представлены как минимум тремя видами пургаториусов, но они малочисленны и представляют меньше пяти процентов от всех индивидов млекопитающих.
А вот 584–691 тысячу лет спустя границы эр к пургаториусам добавляется как минимум ещё один их родственник – Pandemonium dis (кстати, отличное родовое название!). Хотя приматы по-прежнему отстают по видовому разнообразию от прочих зверей, но самое важное – с этого момента приматы надёжно обгоняют предков копытных в численности, составляя четверть от всех индивидов млекопитающих.
855–1148 тысяч лет после вымирания динозавров число видов приматов вырастает совсем незначительно, безнадёжно отставая от копытных, зато один только примат Paromomys farrandi представляет 56 % от всех индивидов, при том, что все копытные составляют меньше 12 %!
И лишь спустя миллион с лишним лет после начала нашей эры копытные достигают 29 % от числа индивидов.
Понятно, что среди плезиадаписовых быстро пошла адаптивная радиация, то есть взрывное увеличение разнообразия. Каких только версий не появилось!
Одни – Plesiadapidae – выросли в размерах и расселились от Европы и Северной Америки до Пакистана и Китая. Образцовый Plesiadapis tricuspidens был довольно тяжеловесным белкоподобным зверем весом от двух до пяти килограмм. Между прочим, это были самые большие и одновременно самые многочисленные звери в своих фаунах! Во французском местонахождении Мена сохранились отпечатки двух тушек, благодаря чему мы знаем, что у Plesiadapis insignis были пушистые хвостики. В питании плезиадаписы сделали ставку на растительноядность, от ранних форм к поздним в рационе увеличивалась доля фруктов и, особенно, листьев. Подобно многим другим растительноядным зверям, зубная система плезиадаписов специализировалась: резцы превратились в грызущие долота, клыки, а иногда и вторые резцы исчезали, моляры же стали жевательными. Впрочем, резцы так и не обрели способности к постоянному росту и самозатачиванию, как у грызунов, отчего, вероятно, плезиадаписы в итоге и проиграли эволюционную гонку. Да и коротенькие лапки с когтями были приспособлены больше для неторопливого лазания, чем для прыгания, а с повлением новых, более бодрых древесных хищников, такой способ передвижения оказался проигрышным.
Saxonellidae и Carpolestidae имели похожее строение зубов, но гораздо более мелкие размеры – с мышь или крысу. В развитие специализации карполестиды обрели высокие премоляры с пильчатым краем, которым резали волокнистые плоды, орехи и стебли. В отличие от плезиадапид, на большом пальце ноги карполестиды уже имели настоящий ноготь – это было прогрессивно, но, впрочем, не спасло их от вымирания.
Paromomyidae стали фруктоядами, отчего их нижние резцы удлинялись и истончались, а моляры становились всё более квадратными и уплощёнными. По некоторым признакам паромомииды были похожи на шерстокрылов, хотя планирующих способностей не обрели.
Иные группы пошли по пути миниатюризации. Picrodontidae стали питаться не только фруктами, но и пыльцой и древесным соком; например, у Zanycteris череп резко сужен спереди, как у современных мышиных лемуров Cheirogaleinae и хоботноголовых кускусов Tarsipedidae, сосущих нектар через трубочку своих челюстей. Micromomyidae уменьшились – или не выросли – до 20–30 г, при таких размерах им не оставалось ничего, как ловить насекомых в древесной листве.
Как обычно, наши прямые Великие Предки избежали всех соблазнов и остались максимально генерализованными. Palaechthonidae – Palaechton и Plesiolestes – выглядели крайне невзрачно и даже примитивно: усреднённые пропорции, маленькие глазницы, самые банальные зубы без увеличений и уменьшений, редукций и гипертрофий. Палехтон имел неплохое обоняние и питался в основном насекомыми. Из него или кого-то очень похожего, разве что чуть более древесного, и появились мы.
В конце палеоцена могли возникнуть уже и настоящие приматы, а не какие-то недоделанные приматоморфы. Один из претендентов – китайский Petrolemur brevirostre, известный, впрочем, по одному обломку верхней челюсти. Разные палеонтологи определяли его и как полуобезьяну или представителя Adapidae, и как анагалида Anagalida, и как парнокопытное Artiodactyla, и как хищного родственника копытных из семейства Oxyclaenidae. Из этого разброса мнений понятно, что кандидат он тот ещё. Но не беда, ведь есть ещё китайский же Decoredon anhuiensis, даже ещё более древний, среднепалеоценовый, зато представленный и верхней, и нижней челюстями и куда как более прогрессивный, так что его порой относят уже к омомисовым обезьянам. Правда, некоторые антропологи всё же сомневаются в его приматности, а кое-кто определял его как кондиляртра, так что и тут сомнения остаются. Такая уж беда с примитивными тварями – все они на одно лицо!
Мораль этих процессов проста: приматы были одними из самых первых плацентарных млекопитающих. Они появились где-то на самой границе мезозоя и кайнозоя, то ли чуть раньше, то ли совсем чуть позже; показательно, что даже в самом раннем достоверном изводе есть уже как минимум два вида, что предполагает какую-то предшествующую эволюцию, которую ещё предстоит прояснить. С самого начала приматы заняли специфическую нишу всеядных древолазов, и этим была предопределена наша дальнейшая эволюция. Несмотря на небольшое и медленно росшее разнообразие и вроде бы медленные темпы размножения, приматы быстро стали едва ли не доминирующей по численности особей группой: с самого начала мы стремились захватить планету – и нам таки это удалось!
Оставалось совсем ничего – слезть обратно на землю и стать людьми. И за этим дело не стало, но – уже в другие эпохи.
Граница палеоцена и эоцена вроде бы не была такой уж ужасной. Температуры плавно росли, планета зарастала. Собственно, так бы всё плавно и развивалось, да вот 56 миллионов лет назад не то земля-матушка выделила CO2 больше нужного, не то красно солнышко пыхнуло от души, не то земная ось содрогнулась, да так, что температуры разово подпрыгнули на несколько градусов, а затем упали обратно. Ясно, что такие выкрутасы не могли совсем не сказаться на планктоне, по которому, собственно, и проводится граница эпох. Но изменения были не такими уж и существенными, а во многих разрезах провести внятную границу вообще не удаётся.
Первые десять миллионов лет после позднемеловой катастрофы планета приходила в себя. В мире невзрачных карликов гигантами были разве что черепахи, змеи, крокодилы да некоторые птицы. Масса серых однообразных млекопитающих стремительно занимала освободившиеся экологические ниши, землеройкоподобные зверьки на глазах превращались в мелких и крупных копытных, всеядных и хищников, меняли свои челюсти, зубы и конечности. К концу эпохи возникла уже примерно половина современных отрядов, которые соревновались с массой странных альтернативных версий.
Наши предки на общем фоне выглядели вполне достойно: начав с весьма скромного старта, они быстро набрали разгон и к следующей эпохе подошли во всей готовности. Невзгоды в виде хищников и конкурентов не могли остановить нашего прогресса, а лишь подстёгивали его. Благо, климат способствовал нашему триумфу.
Альтернативы
Как минимум во второй половине мела отличные шансы на захват планеты имели птицы. Динозавры становились всё больше, а самих их становилось всё меньше. Экологические ниши мелких и среднеразмерных наземных существ освобождались. Судя по обилию видов и появлению даже нелетающих форм, птицы неплохо продвигались к занятию доминирующего положения. Но у них были свои проблемы. Большинство из них связано с полётом.
В частности, активный полёт требует максимального облечения веса и мощнейшего обмена веществ. С таким сочетанием очень трудно иметь крупный затратный мозг. Конечно, врановые и попугаи показывают чудеса интеллекта, а в решении отдельных задач превосходят даже многих людей (попробуйте на досуге решить задачки по добыванию вкусняшек из ящиков, предлагаемые птичкам в экспериментах), но ограничения на размер и работу мозга у них слишком жёсткие. С тяжёлой головой слишком далеко не улетишь. Конечно, большой мозг можно нарастить, потеряв способность к полёту, но и тут незадача: нелетающие птицы всегда возникали в каких-то очень специфических условиях. Либо это происходило в полной безопасности на островах, как с дронтами, моа и эпиорнисами, но тогда не было стимула для прокачки интеллекта, состояние которого у додо вошло в поговорку. Либо птицы становились бескрылыми в пограничных экологических нишах типа побережий – гесперорнисы, пингвины и бескрылые гагарки, – но и тут развить разумность проблематично, слишком узкие задачи ставит специфическая среда. Наконец, можно перестать летать и в обычной среде, но придётся стать либо быстрым или скрытным, как патагоптериксы и цесарки, либо большим и способным запинать любого обидчика, как страусы. Но, если никакой враг не страшен, то зачем быть умным?
Очень подвела птиц специализация крыльев: свободные пальцы исчезли, а кинетизм черепа с подвижным надклювьем, каким бы совершенным он ни был, не мог компенсировать отсутствие ручек. Орудийная деятельность птицам не то чтобы совсем заказана – галапагосские вьюрки и новокаледонские вороны пытаются как могут, – но ограничена до предела.
Есть и более специфические проблемы. Суперобмен веществ – это круто, но требует особого двойного дыхания с хитрым строением лёгких и воздушных мешков. У птиц перед вылуплением лёгкие приходят в готовность очень долго: до трёх суток они подсушиваются, амниотическая жидкость всасывается в кровь, в это время эмбрион сначала дышит из воздушной камеры яйца, а потом ещё сутки – наружным воздухом через дырочку в скорлупе. Живорождение с таким дыханием невозможно. Да и беременная птица, долго носящая развивающийся крупный плод в себе, вряд ли смогла бы летать. А запаса питательных веществ в яйце намного не хватает. Одно дело – ограниченный желток в яйце, а совсем другое – несколько месяцев бесперебойного питания через плаценту от мамы, да ещё с гарантированным отведением всего плохого (у мамы, конечно, будет токсикоз, но она большая, справится как-нибудь).
Кроме прочего, птичкам надо очень быстро взрослеть, чтобы начать летать и либо избегать опасностей, либо направляться в тёплые края. От силы пара-другая месяцев – вот и всё детство. А с такими темпами, да с таким мозгом многому не научишься. Вот и приходится всё поведение записывать в виде рефлексов и инстинктов. А это – антитеза разумности, каковая по определению – нелинейность, нестандартность реакций с учётом предыдущего опыта. Конечно, большинство млекопитающих тоже растут очень быстро, и наши предки в некоторые моменты были не таким уж исключением, но у нас, по крайней мере, есть потенциальная возможность растянуть детство и процесс обучения подольше; некоторым удаётся оставаться детёнышами и десятилетиями. Век живи – век учись!
Имеются у птиц и биохимические сложности. Как известно, над хвостом у них есть копчиковая сальная железа, выделяющая смазку. Большинство людей думает, что птички «чистят» себя, на самом деле они клювом мажут на пёрышки сало. Сало же это не простое, а с двойным назначением. Во-первых, оно делает кератин перьев более эластичным. Во-вторых, из него облучением солнечным ультрафиолетом получается витамин D. Вообще-то, витамин должен бы вырабатываться прямо в коже, да вот беда – она практически полностью закрыта перьями. Вот и приходится птицам сначала обмазываться салом, а потом собирать его, прогоркшее, обратно клювом. Понятно, что делать это в больших количествах сложно и затратно, а без витамина D грозит не только рахит, но и проблемы с работой нервной системы.
В общем, птички – это прикольно, но бесперспективно. Первобытные тинаму и куры упустили свой шанс на господство. Впрочем, если считать виды и численность, может быть, не так уж у них всё и запущено. Ведь мы, в конце концов, живём на планете птиц…
Эоцен. 56–33,9 миллиона лет назад: мир-курорт
Оазис кайнозоя, мечта туриста, последняя передышка планеты перед наступающим ледниковым ужасом. Эпоха благословенного сада, полного сочной зелени и фруктов. Время цветущих Гренландии и Антарктиды, тропических Сибири и Европы.
Но это и переломный момент, когда планета окончательно свернула в пучину замерзания, остыв к концу втрое.
Время, когда планету населяли весьма колоритные существа – уже далеко не такие невзрачные, как в палеоцене, но и далеко не похожие на современных. Пора самых дерзких эволюционных экспериментов, появления причудливых тварей, лишь бледное подобие которых осталось нам в наследство.
Эпоха, когда нашим предкам ещё мало что грозило в их древесном раю, но и этап, когда проблемы и опасности начали настигать их с пугающим ускорением, заставляя эволюционировать.
Первая треть эпохи ознаменовалась продолжением потепления и достижением эоценового климатического оптимума, когда температуры оказались в среднем на 14 °C теплее нынешних! Только в кембрии и девоне было так же здорово, да ещё временами, но недолго. Правда, примерно с пятидесяти миллионов лет назад началось похолодание, по большому счёту продолжающееся до самого недавнего времени. Но мороз для эоцена – благодать для наших реалий. Для сравнения, разница между самым холодным климатом конца эоцена и современным такая же, как между самыми суровыми пиками оледенения плейстоцена и современностью. Другими словами, глядя из ледникового периода, мы живём в межледниковье, а глядя из эоцена – в ледниковом периоде.
Особенно приятно, что повышение температуры было, конечно, не строго пропорциональным относительно современных значений в каждом конкретном регионе, то есть если в Гренландии росли тропические леса, это не значит, что на экваторе на солнце плавился металл. Тепло достаточно равномерно разносилось по всей планете, и везде царил мягкий субтропический климат. Огромные площади поросли тропическими лесами, но в это же время развиваются также травяные степи и саванны, первые образцы коих покрывают среднеэоценовые просторы Южной Америки, а также пустыни – в той же Южной Америке и по северо-восточному краю Африки, в будущей Аравии. Парадоксально, но в эоцене же мы встречаем и первые холодолюбивые леса в горах.
География эоцена всё ещё выглядит экзотично, хотя, конечно, уже несколько приблизилась к современной. Все материки и большие острова немножко сдвинулись на север, особенно это видно для Индии, которая к концу эпохи почти уже доплыла до Азии.
Морские экосистемы эоцена процветали. Перешедшие из мелового периода в кайнозой фораминиферы-нуммулиты Nummulitida (или Rotaliida) только наращивали разнообразие и достигли рекордных размеров. Самым большим оказался Nummulites millecaput – до 16 см в диаметре! А ведь речь идёт об одноклеточных существах! Эти «раковинные корненожки» могли быть не только гигантскими, но и многочисленными: египетские пирамиды сделаны из нуммулитового известняка, сформировавшегося 45 млн л. н. из Nummulites gizehensis. Спасибо корненожкам за Чудо Света! Показательно, что несколько линий нуммулитов оборвались на границе эоцена и олигоцена, для них великое вымирание случилось именно в этот момент – замерзание тропическим существам не идёт на пользу.
Шестилучевые кораллы Hexacorallia в эоцене не чересчур благоденствовали, а всю вторую половину эпохи с ходом похолодания рифостроение всё более и более сокращалось, пока не свелось почти к нулю (как обычно, «не всё так однозначно»: чрезвычайные потепления жахали по рифам в целом и кораллам в частности не хуже заморозков; для этих чувствительных созданий вредны любые экстремумы). Как и в случае с нуммулитами, конец эпохи ознаменовался вымиранием целого ряда линий. По большому счёту, конец эоцена оказался для кораллов столь же губительным, что и конец мезозоя!
Моря эоцена, как ни странно, продолжали хранить в себе крайне архаичных головоногих. В это время появляются наутилусы современного рода Nautilus – N. praepompilius из Казахстана и N. cookanum из США, – выглядящие сейчас как привет едва ли не из палеозоя, так как относятся к очень древней группе Nautilida.
Современные наутилусы прекрасны: их круглая полосатая раковина совершенна в своей лаконичности. В отличие от других современных головоногих, наутилусы имеют огромное количество щупалец – до девяти десятков! Щупальца, кстати, являются производной от ноги, другая часть которой превратилась в характерный головной капюшон. У наутилусов четыре жабры в отличие от прочих современных головоногих с их двумя жабрами. Глаз наутилуса – шарообразная колба с отверстием спереди, заполненная водой, – занимает строго промежуточное положение между светочувствительными пятнышками простых моллюсков и сложным глазом осьминога с роговицей, радужкой, хрусталиком и стекловидным телом. Таким образом, в ряду разных моллюсков можно видеть все этапы эволюции глаза, то есть то, что по палеонтологическим данным никак не прослеживается.
Итальянские глубины прорезали и другие реликты – Bayanoteuthis rugifer – одни из последних белемнитов. Этих головоногих не сгубил позднемезозойский кризис, вымерли они уже в середине кайнозоя, не дожив до наших дней совсем малость.
Ясно, что Италии как страны в эоцене не было, так что корректнее говорить не «итальянские глубины», а «глубины западной оконечности океана Тетис на территории нынешней Италии», но тут и далее в угоду краткости и простоты подобные обороты будут сокращены.
Ихтиофауна эоцена похожа и не похожа на современную. С одной стороны, в это время уже появились все основные группы современных рыб, но все они были немножко другими, чуток недоделанными.
Невероятный аквариум предстаёт в среднеэоценовых отложениях Монте Болька в Италии; на некоторых отпечатках тут отлично видна даже окраска! Рыбы-хирурги Tylerichthys nuchalis и карликовые родственники рыбок-клоунов Odonteus pygmaeus сновали среди кораллов и актиний. Ceratoichthys pinnatiformis, Exellia velifer и Eoplatax papilio с их шикарными высокими плавниками парили, словно бабочки, над песком, в который закапывались бесчисленные скаты – длинноносый Narcine molini, круглый Trygon muricata и ромбовидный Promyliobatis gazolae. Караулили свою добычу морские черти Lophius brachysomus и рыбы-клоуны Histionotophorus bassani. Тут же вились угреподобные Proteomyris ventralis и Bolkanguilla brachycephala и совсем уж нитевидный Whitapodus breviculus. Ловили улиток вытянутые губаны Eolabroides szajnochae и Tortonesia esilis, собирали всякий мусор колючие аргусы Scatophagus frontalis. Рассекали толщу вод мечерылы Blochius longirostris с длиннющей шпагой на носу, а испуганные полурылы Hemirhamphus edwardsi с короткой верхней и очень длинной нижней челюстями выпрыгивали в воздух. В зарослях водорослей маскировались рыбы-иглы Urosphen dubia и Fistularioides veronensis, а мимо не спеша дрейфовали закованные в панцирь аракановые кузовки Eolactoria sorbinii с парой длиннющих рогов спереди и парой сзади, а также колючие рыбы-ежи Oiodon tenuispinus. Рыскали и более стандартные ставриды Carangopsis dorsalis, а также бесчисленное множество иных рыб. Показательно, что больше всего было колючих ночных рыб-белок – Tenuicentrum lanceolatum, Eoholocentrum macrocephalum, Myripristis homopterygius и других, – при том, что в современных коралловых сообществах они составляют явное меньшинство. Напротив, на современных рифах резко преобладают помацентровые, тогда как в Болька они были крайне редки.
Тут же в Болька обнаружены первые камбалы, демонстрирующие самое начало появления асимметрии. У Heteronectes chaneti и Amphistium paradoxum (а также чуть более поздней французской A. altum) тело было уже вполне узнаваемой камбальей формы, но ещё не лежачее и без захода спинного плавника на голову, совершенно обычный рот, а правый глаз только-только начал смещаться на левую сторону, так что эти первобытные камбалы левым глазом смотрели немножко вниз, а правым – сильно наверх. Видимо, они вполне активно плавали как все нормальные рыбы, но периодически ложились на дно на бочок отдохнуть, приподнимаясь, чтобы оглядеться и грести дальше. Между прочим, установить такие явления на окаменелых рыбах очень непросто, ведь на отпечатке череп часто бывает перекошен, и понять причину кособокости – тафономическая она или эволюционная – весьма сложно. Но на то и крутые палеоихтиологи: путём сопоставления мелких косточек правой и левой сторон черепа было доказано, что дело именно во врождённой асимметрии, а не посмертной деформации.
Современные Psettodes erumei из Индийского океана и P. belcheri из Западной Африки выглядят как логичное продолжение эоценовых протокамбал и одновременно как недоделанные камбалы: у них по-прежнему спинной плавник не заходит на голову, рот симметричен, левый глаз находится на положенном месте, а правый расположен строго на макушке.
Впечатляющие рыбы эоцена – Clupeopsis straeleni из Бельгии и Monosmilus chureloides из Пакистана могли достигать метра длины, а их челюсти были усажены дюжиной огромных клыкоподобных зубищ. Рыбы стали саблезубыми до того, как это вошло в моду у котиков! Ясно, что раз среди рыб есть страшные хищники, значит, есть и крупная добыча. Между прочим, современные родственники клюпеопсиса – мелкие планктоноядные рыбёшки-анчоусы; так эволюция может превратить грозу морей в скромного собирателя.
Сухопутные членистоногие эоцена прекрасны по-своему. В Северной Америке доживал свой век последний представитель тараканоподобного отряда насекомых Alienoptera – Grant viridifluvius, чрезвычайно похожий на муравья. Очевидно, мимикрия была весьма актуальна в тогдашних джунглях.
А муравьи были достойны того, чтобы ими прикидываться. Самки гигантских североамериканских муравьёв Titanomyrma lubei достигали 5 см, то есть размеров современных колибри! Ужасны были и самцы немецких Formicium giganteum, похожие на огромных жуков. Впрочем, и в жизни этих монстров были свои сложности: на отпечатках древесных листьев в местонахождении Мессель найдены следы челюстей муравьёв-зомби, поражённых паразитическими грибами кордицепсами. Судя по обилию таких отпечатков, для бедных мурашек засилье коварных грибов было огромной проблемой. В балтийском янтаре сохранился бедняга муравей-древоточец Camponotus, из которого в нескольких местах уже пророс жуткий Allocordyceps baltica.
Некоторые грибы из типа аскомицетов Ascomycota совершенно кошмарны. Кордицепс однобокий Ophiocordyceps unilateralis поражает мозг муравья Camponotus leonardi и делает из него зомби: одурманенная букашка заползает на ствол дерева, вцепляется там жвалами, растопыривает лапки и гибнет. Из головы зомби-муравья вырастает плодовое тело гриба и раскидывает споры на пробегающих под деревом других муравьёв. Другие кордицепсы могут уничтожать иных членистоногих, например Cordyceps ignota – пауков, а Cordyceps tuberculata – мотыльков. Конечно, такое жуткое явление не могло быть обойдено вниманием творческих людей, – и вот уже в нескольких компьютерных играх и фильмах грибы делают зомби из людей. Правда, особо впечатлительные могут расслабиться: человеческий мозг слишком сложен для столь однозначной манипуляции, какую производит гриб с муравьём.
Впрочем, это не отменяет микозов. К слову, у аскомицетов, как и у других грибов, контроль начальных стадий клеточного деления обеспечивается белками SBF вирусного происхождения, а ведь вирусы – профессиональные обманщики иммунитета. Это позволяет грибам легко проникать в клетки растений и животных для паразитирования и симбиоза. Лишь у примитивных и рано отделившихся от других грибов хитридиомицетов Chytridiomycota имеются ещё и транскрипционные факторы E2F, такие же, как у растений и животных.
В тех же немецких лесах над цветами маслиновых и сапотовых деревьев жужжали мухи длиннохоботницы Hirmoneura messelense и собирали пыльцу, остатки которой найдены на их отпечатках. Правда, тогда длиннохоботницы были ещё короткохоботными.
Современные мухи длиннохоботницы Nemestrinidae внешне довольно невзрачны, но обладают длинным негнущимся хоботом, которым высасывают нектар из цветов. Необычно их развитие: из тысяч яиц, отложенных в трещины древесной коры, вылупляются личинки, покрытые развесистыми выростами, отчего ветер подхватывает их и разносит куда попало. А попасть они должны бы на другое насекомое – жука или саранчу; при определённом везении личинка мухи пролезает в тело жертвы через дыхальце и злобно поедает несчастного хозяина изнутри.
Были и более благообразные опылители: бабочки Prodryas persephone и Praepapilio colorado ничем принципиально не отличались от современных. А для гораздо более скромной по размерам бабочки из Месселя удалось даже установить цвет крылышек: передние зелёные с синей и коричневой каёмками, а задние – коричневые с синими краями.
Мессель – уникальная сокровищница Германии, отражающая жизнь среднего эоцена 49 миллионов лет назад. Это был райский тропический сад, раскинувшийся по краям вулканической кальдеры вокруг глубокого озера с болотистыми берегами. Но рай был с адскими нотками: периодически из недр озера поднимались ядовитые вулканические газы, травившие всё живое. Остатки растений и животных погружались на дно и, не разлагаясь, превращались в отпечатки фантастического качества.
Чего только тут не найдено: семена, листья, насекомые, рыбы, лягушки, питоны, крокодилы, птицы, самые разнообразные звери! Окаменелости из Месселя поражают своей детализацией: у членистоногих и птиц часто видно окраску, на рептилиях – каждую чешуйку, а на зверях – шерстинку. У многих животных прекрасно сохранились мышцы и содержимое желудка. На планете очень немного мест, где удалось бы найти столь полную летопись целой экосистемы, да ещё столь богатой.
Другое такое же богатое немецкое местонахождение – Гейзельталь. Здешние лягушки, черепахи, змеи, крокодилы, гасторнисы, архаичные растительноядные, приматы и хищники дополняют картину эоценовой жизни.
Амфибии эоцена принципиально не отличаются от современных, но некоторые уникальны своей сохранностью: Thaumastosaurus gezei из Франции представлена «мумией», на которой – конечно, в окаменевшем виде – сохранились не только общие очертания и складки кожи, но и головной, и спинной мозг, и даже глаза!
Другие лягушки интересны своей географией: сорок миллионов лет назад Calyptocephalella квакали на озёрах, покрытых листьями кубышек Notonuphar antarctica и окружённых лесами из буков Nothofagus – на острове Симор около Антарктиды. Показательно, что и родственные лягушки группы Australobatrachia, и буки-нотофагусы в древности и ныне расселены на юге Южной Америки и в Австралии. Получается, сейчас уцелели только края ареалов, а самое интересное происходило в пущах Антарктиды.
Похожее географическое распространение имели некоторые черепахи. Как и положено антиподам, они очень странные. Аргентинская гигантская наземная черепаха Niolamia argentina имела две пары огромных рогов на затылке, закинутых назад, и один на носу; края её панциря были покрыты шипами, а хвост охвачен плотно пригнанными парношипастыми кольцами, отчего тварь была просто удивительно похожа на мелового монгольского анкилозавра Tarchia gigantea! Кстати, в позднем мелу родственная Otwayemys cunicularius жила в Австралии, а Patagoniaemys gasparinae – в Южной Америке; в Монголии же в мелу обитала не родственная, но внешне похожая Mongolochelys efremovi: как обычно, одни и те же проблемы вызывают одинаковые решения. В кайнозое в Австралии и на рядом расположенных островах насчитывалось не менее шести видов.
Впрочем, из подобных географических вывертов строгое деление фаун на северные и южные никак не следует. Например, древнейший и примитивнейший питон Messelopython freyi найден в немецком Месселе – сильно не рядом с ареалом нынешних потомков. Другой пример хитрой зоогеографии – змеи очень примитивного семейства Madtsoiidae: они найдены в Северной Африке, Южной Европе, Южной Азии, Южной Америке и Австралии, то есть в самых разных местах, но вроде бы в целом по периметру моря Тетис. Некоторые представители могли достигать гигантских размеров, а рекордсменом из чемпионов был египетский Gigantophis garstini: по смелым оценкам – 9–11 м или по скромным – 7 м в длину.
Среди ящериц столь же загадочно расселены тейиды Teiidae: в конце мела похожие крокозябры обнаруживаются в Монголии, Европе и Северной Америке, в эоцене – в Европе, а ныне населяют Южную и Центральную Америку. Во Франции жили ядозубы Eurheloderma gallicum, родственники которых ползали и ныне ползают по полупустыням Центральной Америки (а в позднем мелу Gobiderma pulchrum и Estesia mongoliensis корячились в Монголии); по всей Европе жило множество видов игуан-василисков Geiseltaliellus, при том что современные игуаны греются в лучах солнышка в обеих Америках, на Фиджи и Тонга и – неожиданно – Мадагаскаре.
Конечно, не одной географией жива палеогерпетология. Североамериканский варан Saniwa ensidens имел четыре глаза: два обычных и два на основе верхней части промежуточного мозга – пинеального и парапинеального тел. У миноги есть оба этих тела, расположенные прямо под кожей головы; у гаттерии «третий глаз» – парапинеальное тело – выходит из черепа, но всё же тоже лежит под кожей; у людей имеется «третий глаз» – эпифиз, или пинеальное тело, только он совсем не выходит даже за пределы мозговых оболочек. У санивы дополнительные глазики были крохотными, но пронизывали череп. Любопытно, что у более древних и более молодых родственных видов отверстие на макушке только одно; получается, второе появилось только у одного вида на небольшой временной отрезок. Зачем – загадка, но сама возможность формирования нового органа из старых элементов прекрасна.
В некоторых эоценовых фаунах рептилии и, возможно, птицы всё ещё продолжали быть крупнейшими наземными хищниками. Особенно, конечно, отличились крокодилы, в том числе сухопутные и особенно, конечно, в изолированных областях планеты. Так, по Южной Америке рыскали двух-трёхметровые длинноногие и высоко-узкоголовые крокодилы-мезозухии Sebecus icaeorhinus (названные, кстати, в честь египетского бога Себека). А ведь, между прочим, все их родственники вымерли ещё в конце мела! В Северной Америке, Европе, Казахстане и Китае подобную же роль выполняли чуть менее специфические крокодилы Pristichampsus, Boverisuchus и Planocrania.
Зубы Boverisuchus сильно сжаты с боков и тем очень похожи на зубы хищных динозавров теропод, отчего первоначально и были приняты за динозавровые. А ведь из этого вытекало, что часть динозавров успешно пережила меловое вымирание и дожила аж до эоцена!
В продолжение палеоценовой истории продолжали эволюционировать нелетающие страусоподобные. Особенно показателен немецкий лесной Palaeotis weigelti: метрового роста, со сравнительно большими крыльями, с узким, как у казуаров, клювом. По Швейцарии вышагивали Eleutherornis helveticus, по Франции – Galligeranoides boriensis, по Италии – Palaeogrus princeps, а по Египту – Eremopezus eocaenus (и, возможно, отдельный род и вид Stromeria fajumensis), ещё сохранявший первый – задний – палец на стопе.
Происхождение страусов – по сию пору нерешённый вопрос. С одной стороны, они вроде бы выглядят очень примитивно, с другой – достоверные и несомненные страусы появляются в палеонтологической летописи весьма поздно. Выдвигалась масса гипотез их возникновения. Интригующе выглядит версия, согласно которой страусы – прямые потомки динозавров и вообще никогда не летали; действительно, на их грудине нет ни малейших следов киля, но отчего бы ему и не пропасть вторично. По другой трактовке, предками страусов могли быть эоценовые североамериканские журавлеподобные Geranoididae типа Geranoides jepseni (который может быть синонимом Palaeophasianus meleagroides). По ещё одному предположению, их предки – Eogruidae типа Eogrus из позднего эоцена Монголии, Китая и Казахстана. А может быть, это были олигоценовые и миоценовые Ergilornithidae типа Ergilornis и Amphipelargus из Азии и Европы, которые сами могут быть всё теми же Eogruidae. Почти все эти и многие другие птицы известны по фрагментам костей ног и, иногда, крыльев. Два ярких признака, выделяющих страусов – строение нёба и редукция внутреннего пальца на стопе, определимы с большим трудом. Черепа птиц вообще редко сохраняются, а палеогнатное нёбо могло возникнуть несколько раз из неогнатного через хитрые неотенические перестройки (филэмбриогенез рулит!). Исчезновение третьего пальца тоже могло происходить независимо в разных линиях. Сравнивать разные окаменелости разных форм крайне сложно, так что число догадок о филогенетических взаимоотношениях пташек больше, чем число палеоорнитологов, которые их выдвигают.
Как итог, по морфологии выходит, что страусы могли бы включаться в журавлеобразных Gruiformes, как бы странно это ни звучало. Правда, молекулярные данные эту странность исключают, вынося страусов и тинаму в отдельную эволюционную ветку от всех птиц, весьма далеко от журавлей. В любом случае факт остаётся фактом: древнейшие нормальные страусы Struthio coppensi известны лишь из раннего миоцена Намибии.
Вообще, гигантские нелетающие птицы по-прежнему были в моде. Перуанские пингвины Icadyptes salasi достигали высоты 1,5 м; между прочим, жили они совсем недалеко от экватора, да ещё и в эпоху климатического оптимума (а найдены так и вовсе в пустыне), так что образ пингвина как холодолюбивой птицы – явный перекос нашего современного восприятия. Anthropornis grandis и Anthropornis nordenskjoldi (ростом, между прочим, 1,5–1,8 м и весом от 90 до 135 кг!) из Антарктиды могли бы восстановить реноме, если бы Антарктида сама не была в то время тропическим садом.
Современные пингвины тоже далеко не всегда населяют снежно-леденелые области. Рекордсмены, конечно, галапагосские пингвины Spheniscus mendiculus, живущие строго на экваторе. Практически до той же широты дотягивают пингвины Гумбольдта Spheniscus humboldti, населяющие почти всё тихоокеанское побережье Южной Америки. Очковые пингвины Spheniscus demersus в Южной Африке и малые пингвины Eudyptula minor в Южной Австралии и Новой Зеландии тоже явно не могут быть зачислены в число антарктических жителей. Они, хотя и держатся холодных течений, делают это не из любви к морозу, а лишь потому, что в холодной воде больше планктона и, стало быть, рыбы.

Gastornis
В Евразии и Северной Америке всё шире расселялись гасторнисы: в Месселе и Гейзельтале – Gastornis geiselensis и то ли тот же, то ли другой вид G. sarasini; G. giganteus (он же Diatryma) добрались до Северной Америки, а G. xichuanensis – до Китая. Между прочим, это значит, что подобных птах можно искать и в нашей стране, ведь миграции могли идти либо через Берингию, либо через моря, отделявшие Европу от Азии. Эти чудища известны не только по костям: во Франции найдены следы здоровенных ног и скорлупа огромных – 10–24 см – яиц, а в формации Грин Ривер – отпечаток гигантского пера. Поскольку костей других настолько больших птиц из тех же и близких отложений неизвестно, следы, яйца и перья, скорее всего, принадлежали именно гасторнисам. В конце эоцена гасторнисы вымерли, не сумев приспособиться к жизни на открытых пространствах.
В 1923 году были описаны нитевидные отпечатки из формации Грин Ривер – длиной более 20 см. Был сделан вывод, что это перья гигантской птицы Diatryma fihifera. После этого на всех реконструкциях гасторнисы-диатримы представали наподобие гигантских киви или молотоголовых казуаров, покрытых длинными волосовидными перьями. Однако при ближайшем рассмотрении выяснилось, что нити из Грин Ривер – на самом деле отпечатки растений.
Формация Грин Ривер – огромная толща эоценовых отложений, тянущаяся вдоль реки Грин аж по трём штатам – Колорадо, Вайомингу и Юте. Тончайшие отложения формировались на дне цепи озёр каждый год при чередовании сухих и влажных сезонов как минимум шесть миллионов лет подряд! За это время часть озёр была солёной, часть – пресноводной. Вокруг озёр росли леса из платанов Platanus wyomingensis и веерных пальм Sabalites powellii, в которых жили самые разнообразные существа. В слоях формации сохранились идеальные отпечатки многочисленных рыб, ящериц, крокодилов, птиц и зверей. Между прочим, тут же до сих пор высятся гигантские строматолиты – каменные «пни» до 5 м высотой!
Если слава гасторнисов как хищных птиц в последнее время оказалась незаслуженной, того же нельзя сказать про кариам. Древнейшие представители – длинноногие, крепкоклювые и уже тогда нелетающие Bathornis grallator (они же Neocathartes) появились в середине эоцена Северной Америки, чуть позже Strigogyps sapea и S. robustus гоняли мелкое зверьё в Европе, современные же потомки – плохо летающие Cariama cristata и Chunga burmeisteri – благополучно ловят мышей и ящериц в южноамериканских пампасах. Показательно, что конвергентно в Африке ту же нишу заняли птицы-секретари Sagittarius serpentarius, а по всей планете похожую – аисты и журавли. Конечно, все эти птицы не строгие хищники, а всеядны – могут есть и падаль, и семена, а потому чудесным образом могут рассматриваться и как экологический аналог нас – людей.
Нормальные летающие птицы эоцена известны достаточно неплохо, в значительной степени благодаря шикарным отпечаткам из Месселя. Кого тут только нет! Крошечный зимородок-щурка Quasisyndactylus longibrachis и пёстрый удод Messelirrisor grandis, кукушкоподобный Pumiliornis tessellatus с цветочной пыльцой в желудке и острокрылый стриж Scaniacypselus wardi, длинноногий фламинго Juncitarsus merkeli и сова Messelastur gratulator, очень похожая на сокола, исполинские козодои Paraprefica major и P. kelleri. Все они вроде бы и похожи на своих родственников-потомков, а вроде бы и отличаются в мелочах и частностях.
Некоторые отпечатки в Месселе столь качественны, что позволяют оценить цвет перьев по меланосомам: зимородок-щурка Eocoracias brachyptera был чудесной синей птичкой с чёрными крылышками и хвостиком.
В числе прочих пернатых эоцен подарил нам попугаев. Найти их в Индии – Vastanavis eocaena и V. cambayensis – не так удивительно, а вот в немецком Месселе – Psittacopes lepidus, Англии – Palaeopsittacus georgei – и Франции – Quercypsitta sudrei и Q. ivani – всё же неожиданно. Между прочим, эоценовые попугаи всё ещё имели довольно-таки непопугайный – тонкий и длинный – клюв, а их лапки были ещё полу-зигодактильными, то есть два пальца смотрели вперёд, один назад, а один поворачивался. С другими птицами география устраивает ещё более заковыристые выкрутасы: немецкие лягушкороты Masilopodargus longipes из Месселя родственны вайомингским Fluvioviridavis platyrhamphus из Грин Ривер, при том что современные живут от Южной Индии до Австралии и Тасмании.
Млекопитающие эоцена чудесны: они уже не столь однообразны, как палеоценовые, но ещё далеко не так похожи на современных, чтобы быть совсем уж привычными и банальными.
До конца эпохи протянули многобугорчатые, последними были североамериканские Ectypodus lovei, чьи нижние зубы достигли в своём роде апофеоза: огромный резец спереди, огромный гребнистый премоляр в середине и два бугристых моляра в конце. На границе олигоцена они вымерли из-за конкуренции с грызунами и приматами и появления новых хищников. В Аргентине коротко-широкомордые Groeberia minoprioi – родичи многобугорчатых из группы Gondwanatheria – выжили и дали потомков в последующие эпохи.
Сумчатые Marsupialia продолжали процветать в самых разных местах: многочисленные виды Peratherium в Европе и Африке (они дожили аж до миоцена), Asiadidelphis в Казахстане. Особое внимание обращает на себя Anatoliadelphys maasae – очень примитивное хищное сумчатое из Турции. Эта зверюшка весила 3–4 кг – как кошка или сумчатая куница; на последнюю анатолиядельфис был, видимо, похож и по способу передвижения – частично наземный, частично древесно-лазающий довольно бодрый хищник-падальщик. Судя по широким третьим премолярам, анатолиядельфисы любили грызть что-то очень твёрдое, видимо, кости. Из всех евразийских сумчатых это был самый крупный хищник. Впрочем, Турция в тот момент (и задолго до) была островом – этакой Австралией в миниатюре, так что тут могли реализоваться всякие странные экологические связи.
Гораздо более суровые хищники получились из сумчатых в Южной Америке, где сформировались Sparassodonta, или Borhyaenoidea, в лице первой группы Proborhyaenidae. Начав, как и все, с опоссума размером с куницу, они быстро превратились в двухметровых тридцатикилограммовых Callistoe vincei. Череп каллисто был хоть и узкий, но очень мощный, а главное – с огромными клыками на верхней и нижней челюстях, причем нижние были едва ли не больше верхних. Самое замечательное, что клыки не имели эмали и могли неограниченно или почти неограниченно расти, подобно резцам грызунов. Мощнейшие лапы с огромными когтями явно были приспособлены к рытью. Видимо, главной добычей каллисто были норные животные типа броненосцев – тогда и постояннорастущие клыки обретают особый смысл. Порой диггерский пыл каллисто зашкаливал: на типовом скелете локтевая и лучевая кости имеют следы заросшего перелома. Arminiheringia auceta, A. cultrata и A. contigua были массивнее – до 80 кг – и с несколько иными зубами: нижние клыки были растопырены, а заклыковые зубы туповаты, что выдаёт падальщика и всеядного зверя.
Как и прочие существа, сумчатые любили путешествовать: эоценовые южноамериканские опоссумы Zeusdelphys complicatus более родственны меловым североамериканским, а не местным, с которыми они, впрочем, конвергентно очень схожи. Видимо, это говорит о дополнительных миграциях.
Продолжались и прогулки по Антарктиде, где по острову Симор скакали Antarctodolops dailyi и A. mesetaense, представители специфического отряда Polydolopimorphia.
В палеоцене в Австралии млекопитающие неизвестны, но в эоцене этот недостаток был восполнен. Путь сумчатых из Южной Америки в Австралию прослеживается по зубам соневидных опоссумов Microbiotheria, которые те оставляли, двигаясь по дремучим антарктическим лесам, как заблудившиеся детишки из сказок, бросавшие хлебные крошки, чтобы найти обратный путь. Изолированные зубы подобных тварюшек найдены в среднеэоценовых отложениях всё того же западноантарктического острова Симор – Marambiotherium glacialis и Woodburnodon casei – и раннеэоценовых слоях фауны тингамарра в северо-восточной Австралии, где древнейшее сумчатое Djarthia murgonensis выглядит чрезвычайно генерализованно, но ближе всего именно к соневидным опоссумам. Также очень наглядно существование вида Chulpasia mattaueri в Перу и вида того же рода Ch. jimthorselli в раннеэоценовой Австралии; правда, определение австралийской находки вызывает сомнения, но как минимум сумчатость её несомненна.
Кстати, в последующем сумчатые переправились из Австралии в Индонезию, почти завершив удивительное кругосветное путешествие длиной в миллионы лет, ведь древнейшее сумчатое известно из Китая!
В популярных книгах по биологии часто звучит, что Австралия – континент, заселённый только сумчатыми, дескать, из плацентарных сюда смогли долететь лишь летучие мыши да доплыть люди с собаками-динго. На самом же деле, в Австралии и на окружающих архипелагах живёт с полсотни эндемичных видов грызунов, попавших сюда совершенно независимо от людей.
Плацентарные эоцена демонстрируют наиширочайший спектр существ: от совершенно банальных до почти-фантастических.
Продвинутые цимолесты Pantolesta дали несколько разных линий. Одна из них – выдроподобная версия, представленная множеством видов североамериканских Palaeosinopa и Pantolestes, причём палеосинопы появились ещё в конце палеоцена в Северной Америке, а потом расселились и в Европу, где их потомки – Buxolestes piscator и B. minor найдены в разных местах, но лучшие отпечатки, конечно, в Месселе. Это были вытянутые приземистые зверьки почти метровой длины на коротких сильных гребных лапах с пальцами, как у бобра, с мощным уплощённым хвостом, развитыми спинными мышцами и мощными молярами для разжёвывания моллюсков и рыб. Внутри желудка на некоторых отпечатках сохранились останки рыб, так что образ жизни вырисовывается весьма очевидный.
Совсем другая линия – Ptolemaiida – не то часть пантолестов, не то самостоятельный отряд, не то часть афротериев, родственная трубкозубам (первоначально же они были описаны вообще как приматы). Они известны с границы эоцена и олигоцена египетского Файюма и Кении, а последние в Кении дотянули до миоцена. Лучше всего сохранился череп Ptolemaia lyonsi. Это были довольно крупные – с большую собаку – животные с плоской узкой вытянутой головой. Зубы птолемайид высококоронковые гипсодонтные, с тонкой эмалью, но с округлыми бугорками, как у приматов, а не складками эмали, как у копытных. Большие клыки сочетались с огромными третьими и четвёртыми премолярами и маленькими молярами. Челюсти хорошо двигались по вертикали, но плохо жевали. Не исключено, что такими зубами птолемайи разгрызали насекомых или моллюсков.
Среди насекомоядных в начале эоцена появляются первые достоверные ежи – китайские Changlelestes dissetiformis, а уже на границе с олигоценом – предки кротов – европейские Amphidozotherium cayluxi и Eotalpa anglica. Если вдуматься, и ежи с их иголками, и кроты с их лапами-лопатами и потерей зрения – крайне необычные твари, а кажутся привычными только потому, что мы знаем их с детства.
Среди прочих раннеэоценовых насекомоядных выделяется Batodonoides vanhouteni – при весе 0,93–1,82 г, в среднем 1,3 г, это было одно из самых маленьких млекопитающих всех времён! Очевидно, что такие микромасштабы батодоноидес мог иметь только в сверхтёплом климате и сверхобилии насекомых.
Некоторые современные теплокровные приближаются к рекорду батодоноидеса, но всё же в полтора-два раза крупнее. Из млекопитающих это этрусская землеройка Suncus etruscus длиной 3–4,5 см (плюс хвост 3,5 см) и весом от 1,2 до 2,7 грамма, а также свиноносая летучая мышь Craseonycteris thonglongyai из Таиланда и Бирмы – 2,9–3,3 см и 1,5–3 г. Среди птиц свои рекордсмены: ямайская колибри карликовая пчёлка Mellisuga minima имеет длину с клювом 6–7 см, самцы весят 2–2,4 г, максимум 2,7 г, а самки весом 3,6 г несут яйца 1 см длины и 0,375 г весом; у кубинских колибри-пчёлок Mellisuga helenae самцы длиной 5,51 см и весом 1,6–1,95 г, самки – 6,12 см и 2,6 г, яйца длиной 6,35–11,4 мм.
Вообще, наземной и почвенной еды – червей, муравьёв, термитов, жуков, личинок, клубней, луковиц – в эоцене более чем хватало, так что не странно, что параллельно возникло несколько похожих линий ценителей подобного меню.
В Южной Америке таковыми стали неполнозубые Xenarthra. Первые известны из начала эпохи по щиткам панциря Riostegotherium и другим подобным из Бразилии и Аргентины, но толком про них ничего не известно. Впрочем, в том же раннем эоцене жил и хорошо сохранившийся броненосец Utaetus buccatus, уже вполне узнаваемый, но ещё сохранявший эмаль на редких колышковидных зубах, чего у более поздних и современных броненосцев уже нет.
Положение неполнозубых среди млекопитающих – великая загадка, слишком уж они специфичны. Существует три основные версии: в первой неполнозубые выделяются за пределы всех прочих плацентарных, которые тогда обозначаются как Epitheria; во второй афротерии всё же отстоят от других плацентарных больше, чем неполнозубые, в этом случае все, кроме афротериев и неполнозубых, объединяются в Exafroplacentalia (или Notolegia); в третьей неполнозубые с афротериями объединяются в Atlantogenata и противопоставляются Boreoeutheria.
Неполнозубые не просто так получили своё название, у них действительно не всё в порядке с зубами. Ленивцы и броненосцы не имеют ни резцов, ни клыков (только у двупалых ленивцев хорошо развиты клыки), а заклыковые зубы у них очень примитивные – одинаковые, колышковидные, без корней и эмали. Примитивное ли это состояние или же вторичное – неочевидно. У муравьедов зубов вообще нет. Скуловые дуги нормальные у броненосцев, хитро преобразованы в странные незамкнутые закорючки у ленивцев, а у муравьедов вообще редуцированы. Из-за всего этого неполнозубые фактически не могут жевать, да и кусают так себе, а потому имеют специальный мускульный отдел желудка. Чрезвычайно специализированы лапы и позвонки. В частности, на лопатке есть дополнительная ость, клювовидный отросток лопатки (формирующийся как самостоятельный коракоид и поздно прирастающий к лопатке) сливается с плечевым, когти на пальцах огромные, а число пальцев может сокращаться до двух или трёх. Число шейных позвонков у ленивцев отличается от стандартных для млекопитающих семи – от пяти до девяти, причём меняется индивидуально, грудных рекордное число – до 24–25, на позвонках есть дополнительные суставные поверхности, а седалищная кость таза сочленяется с хвостовыми позвонками. Кожа склонна к образованию ороговевших пластинок, которые у ископаемых гигантских ленивцев были подкожной бронёй, у муравьедов имеются в хвосте, а у броненосцев развиваются в полноценный панцирь. Уникален язык муравьеда – больше полуметра длиной, крепящийся аж к грудине, до которой достают и слюнные железы. Всё это вместе взятое говорит о долгой независимой эволюции группы, видимо, уходящей корнями в мезозой.
Некоторые существа зависают между отрядами, даже несмотря на хорошую сохранность образцов. Eurotamandua joresi из Месселя известен по отпечатку целой тушки, на которой видно все детали, но всё равно специалисты спорят, относится ли он к муравьедам из отряда неполнозубых, к панголинам или к особой группе Afredentata в составе афротериев. Внешне евротамандуа действительно сильно похож на тамандуа – современного муравьеда из Южной и Центральной Америки: вытянутая в трубочку беззубая морда, короткие мощные лапы с огромными когтями, цепкий хвост. Многим кажется проблемой география, ведь муравьеды во все времена считались сугубо южноамериканской группой, но среди самых разных животных эоцена полно примеров географических загадок. Для других более показательно отсутствие у евротамандуа чешуи, а ведь роговая чешуя, покрывающая тело как шишку, это едва ли не самый яркий признак современных панголинов.
Гораздо надёжнее относится к панголинам Euromanis krebsi и Eomanis waldi всё из того же Месселя. Спина эоманиса уже точно была покрыта чешуёй, хотя ноги и хвост пока ещё не забронировались. Между прочим, благодаря идеальной сохранности содержимого желудка мы знаем, что эоманисы ели не только насекомых, но и растения.
Получается, что в Европе экологическая ниша расковыривателей муравейников и термитников была занята панголинами Pholidota. Тем более здорово, что они быстро пробрались и в Северную Америку, где по деревьям лазали цепкохвостые панголины Patriomanis americana. Вероятно, ещё позже они обосновались в Африке и Азии, где и живут до сих пор.
Непривычнее для нашего северного взгляда Palaeanodonta – родственники панголинов или, возможно, неполнозубых (при том, что сами панголины и неполнозубые друг другу сильно неродственны). Палеанодонты были коротконогими и очень вытянутыми зверьками в полметра – метр длиной (причём половину составлял хвост) с весьма странным низким продолговатым плоским черепом. Зубы североамериканских палеанодонтов прогрессивно уменьшались: если у Alocodontulum atopum на нижней челюсти имелись резцы, клыки, три премоляра и три достаточно развитых моляра, то у Metacheiromys marshi и M. dasypus на верхней челюсти имелся лишь один клык и один редуцированный заклыковый зуб, а на нижней – один резец, клык и пара крошечных однокорневых заклыковых зубов. Судя по мощным лапам со здоровенными широкими когтями, палеанодонты выкапывали себе пищу из земли, а судя по пропаже зубов, пища не требовала пережёвывания. Тем более странно, что клыки-то у палеанодонтов довольно большие; возможно, они больше использовались для внутривидовых демонстраций, чем для питания.
Первое описание Metacheiromys было сделано по фрагментам скелета, в который по недоразумению были включены части от каких-то приматов. Ошибочно посчитали, что конечности метахейромиса похожи на руконожковые, за что он и получил свое название (Cheiromys – один из её старых синонимов) и был расценён как примат. Просто удивительно, насколько метахейромис в действительности не похож ни на приматов в целом, ни на руконожку в частности – и внешне, и по образу жизни!
Спасаясь от изменений климата конца эоцена, некоторые палеанодонты изобретали экзотические адаптации. Североамериканские Xenocranium pileorivale и Epoicotherium unicum стали подземными копателями – аналогами кротов и златокротов. Их головы стали короткими, с широкими вогнутыми лопатообразными носами, зубы упростились и превратились в редкие колышки, хотя клыки остались большими и толстыми. Слуховые части невероятно увеличились и превратились в широко расставленные в стороны пузыри, слуховые косточки утяжелились и увеличились, обеспечивая низкочастотный подземный слух. Для отгребания земли шейные позвонки с третьего по пятый срослись, плечевая кость превратилась в странную раскоряченную загогулину, в которой узнать плечевую кость – уже непростая задача; столь же искажёнными оказались и кости предплечья, а кисть стала загребущей когтистой лопатой-ковшом. Показательно, что в тех же слоях часты и кости амфисбены Spathorhynchus natronicus.
Родственный ксенокраниуму и эпойкотерию раннеэоценовый североамериканский Tubulodon выдвигался в качестве кандидата на предка трубкозубов, так как его зубы имеют дентиновые трубочки, но эта связь вряд ли оправданна, так как трубочки у него не той системы.
Травоядные эоцена весьма разнообразны. Крупнейшими были Pantodonta. В начале эпохи клыкастые тапироподобные Coryphodon eocaenus немножко уменьшились по сравнению с позднепалеоценовыми, но потом снова достигли размеров коровы и веса 700 кг у позднейшего вида C. lobatus; правда, судьбинушка у них не заладилась, и в середине эоцена корифодоны вымерли. Корифодоны жили в жарких тропических болотах, тем более примечательно обнаружение вида C. pisuqti на острове Элсмир, расположенном рядом с северной Гренландией. Эх, хорошо было в эоцене! На границе палеоцена и эоцена корифодоны проникли из Северной Америки в Монголию, Казахстан и Китай, а чуть позже – и в Европу. Последний представитель группы – Hypercoryphodon thomsoni – дожил до самого конца эоцена или даже олигоцена во Внутренней Монголии. Он достиг размера носорога. Череп гиперкорифодона выглядит средним между свиным и носорожьим, только с узкими скуловыми дугами. Колоритен и Eudinoceras mongoliensis со свиноподобной нижней челюстью и торчащими в стороны нижними клыками.

Coryphodon eocaenus
Конкуренты пантодонтов тоже не стояли на месте. Tillodontia тоже только в эоцене попали в Европу. В своём роде апофеозом развития группы стал Trogosus – крупный растительноядный зверь, отдалённо похожий на хвостатого медведя весом в полтораста килограмм. Тридцатисантиметровый череп с мощнейшими челюстями впечатляет, но, судя по огромным постояннорастущим резцам и стёртым молярам, трогозусы питались какой-то грубой растительной пищей типа корневищ.
Среди Taeniodonta самым крупным был Stylinodon mirus – весом в 80 кг, а его родич S. inexplicatus – вдвое меньше. Мордой стилинодоны были более чем импозантны: огромнейшая нижняя челюсть, которой позавидовал бы любой супермен, огромнейшие боковые резцы и особенно клыки, росшие на протяжении всей жизни, подобно грызунячьим резцам, огромнейшие когти на коренастых лапищах, причём передние были заметно сильнее задних. Видимо, эти монстры выкапывали своими граблями грубоволокнистые корни и клубни и грызли их своими зубищами.
Огромные грызущие звери были очень колоритны, но не могли противостоять новой группе – грызунам Rodentia. Отличный грызущий аппарат, многочисленность и быстрое размножение сделали грызунов сверхуспешными; неспроста нынче это самый многочисленный и по количеству видов, и по количеству особей отряд млекопитающих. Эоценовых грызунов уже очень много. Судя по костям лапок крайне примитивных грызунов Tamquammys robustus и T. wilsoni, двигались они примерно как крысы, может, чуть лучше лазая по ветвям. Особенно удались многочисленные виды белкоподобных Paramys, расселившиеся и по Северной Америке, и по Европе, и по Азии, а также чуть более поздних и более крупных североамериканских Ischyromys. Иширомису родственен и древесный цепкохвостый «дикобраз» Ailuravus macrurus из Месселя.
Очень показательна эволюция мозга грызунов, она шла ровно в тех же направлениях, что и эволюция мозга ранних приматов: мозжечок и особенно полушария конечного мозга увеличивались, а обонятельные доли уменьшались. На первых порах парамисы имели даже относительно больший размер головного мозга, чем синхронные ранние приматоморфы! Правда, процент развития новой коры конечного мозга у грызунов отставал, а темпы прогресса у наших предков были куда бодрее, так что мы вырвались вперёд и стали умничками-разумничками, а белки стали белками.
Как и приматы, первые грызуны освоили древесный образ жизни и питание растениями, но сделали это по-своему. Приматы научились далеко скакать, а грызуны предпочитают перелезать с ветки на ветку без длинных прыжков. Приматы налегли на сочные фрукты, а грызуны – на сухие семена и орехи. Также приматы оказались более-менее универсалами, тогда как грызуны очень специализированно приспосабливаются к каждой конкретной экологической нише с образованием новых видов. Те приматы, которые пытались пойти по грызунячьему пути – несколько линий плезиадаписовых с огромными резцами, – в итоге вымерли, так как, во-первых, не смогли развить столь совершенную жевательную мускулатуру, а во-вторых, вероятно, медленнее плодились.
В самом конце эоцена возникли архаичные бобры Agnotocastor и Anchitheriomys: с этого момента судьба водоёмов была предопределена; впрочем, многие потомки первых бобров стали наземными и даже норными, просто до наших дней дошли лишь любители плавания.
Прорыв грызуны совершили и в географическом смысле: ещё в середине эоцена, около 41 миллиона лет назад в Южной Америке появляются первые кавиоморфные грызуны Caviomorpha – всем известна морская свинка, а кто-то знает и шиншиллу, дегу и капибару. Первые представители – Cachiyacuy contamanensis, C. kummeli и Canaanimys maquiensis – найдены в Перу, а ведь ближайшие более-менее синхронные родственники жили в Африке, Южной и Восточной Азии, но не в Северной Америке, которая к тому же была отделена от Южной проливом. Стало быть, грызуны преодолели Атлантический океан напрямую! И стало быть, до Перу протошиншиллам надо было ещё добираться поперёк всего континента.
Один из ближайших современных внеамериканских родственников кавиоморфных грызунов – Laonastes aenigmamus – неожиданно нашёлся в Лаосе (причём сначала на мясном рынке, потом в совиных погадках и лишь после целенаправленных поисков – вживе). Эта серая «крысобелка» оказалась достойной выделения в самостоятельное семейство Laonastidae или включения в семейство Diatomyidae, известное по нескольким ископаемым родам. Ясно, что заселение Южной Америки не шло из Юго-Восточной Азии через Тихий океан и даже не через Северную Америку, просто потомки неких африканских предков разбрелись в разные стороны: одни поплыли на запад, другие побежали на восток. В самой Африке тоже остались родичи – землекопы Bathyergidae и скальные крысы Petromus typicus.
Своим путём двинулись другие представители Mixodontia из семейства Mimotonidae – монгольские Gomphos elkema и киргизские Anatolmylus rozhdestuenskii: они и их родственники стоят в основании зайцеобразных Lagomorpha. В отличие от грызунов, они научились быстрее бегать, а также питаться корой и травой. По сравнению с грызунами, зайцеобразные стали заметно более универсальными: число их видов гораздо меньше, а экологическая пластичность некоторых видов – гораздо больше. Правда, грызунов такое количество и разнообразие, что любые масштабные обобщения в любом случае натыкаются на обилие исключений.
Существует два крайних полюса экологической пластичности – терпимости – устойчивости живых существ: стенобионты имеют очень жёсткие требования к среде и живут в узком диапазоне условий, а эврибионты – универсалы, способны жить где попало и есть что придётся. Отличные примеры стенобионтов – панды, коалы и муравьеды, а эврибионтов – волки и люди.
Широкое распространение грызунов, вместе с приматами, привело к сокращению числа, а потом и вымиранию многобугорчатых Multituberculata. Как минимум с середины эоцена грызуны начинают отвоёвывать ресурсы и у копытных; по крайней мере, грызунов постоянно становится всё больше, а копытных – относительно меньше.
В преддверии новых времён возникали и ещё более новаторские группы. Чрезвычайно успешными, без сомнения, оказались рукокрылые Chiroptera, ведь они в итоге стали вторым после грызунов по числу видов отрядом млекопитающих. Кто породил летучих мышей – пока неизвестно, видимо, это были какие-то палеоценовые насекомоядные типа цимолестов; прямые предки наверняка давно уже найдены, но таятся в обилии трудноразличимых палеоценовых зубов. Даже у стопроцентных среднеэоценовых летучих мышей зубы сложно отличить от землеройковых, что уж говорить о пращурах.
Древнейшая раннеэоценовая летучая мышь Australonycteris clarkae найдена в Австралии; очевидно, что туда надо было ещё долететь, так как в самой Австралии возникнуть было вроде как не из кого. При этом она ещё и не самая примитивная и даже – правда, сугубо предположительно – уже могла иметь способности к эхолокации. К сожалению, от австралониктериса сохранилось совсем немного. Зато практически идеально сохранились скелетики едва более поздних Onychonycteris finneyi из Вайоминга – самых примитивных летучих мышек. В частности, у онихониктериса сохранялсь коготочки на всех пяти пальцах ручки, не было эхолокации, зато был длинный хвостик. Однако все прочие летучемышьи особенности выражены практически полностью, крылья большие и полноценные, хотя некоторые аэродинамические нюансы и отличаются от современных. После летучих мышей становилось только больше и больше: Icaronycteris index, I. menui и I. sigei из Северной Америки, Франции и Индии, с пяток видов Archaeonycteris из Европы и Индии – и прочая, и прочая.
Рукокрылые – идеальный пример сверхбыстрой эволюции в начале группы, последующей остановки прогресса на большом таксономическом уровне после достижения оптимума и интенсивного видообразования во все времена. В палеоцене мы не видим летучих мышей ни в каком виде, в начале эоцена они возникают практически в готовом виде, а потом у них ничего принципиально не меняется, но виды с частными видоизменениями возникают в огромном количестве. Тут-то бы и закричать всяким креационистам о сотворении, но вот только такая модель эволюции многократно повторялась и до, и после в самых разных группах. То же самое было и с акулами, и с двоякодышащими рыбами, и с черепахами, и с птицами, и с бесчисленным количеством самых разных тварей, причём, чем специфичнее образ жизни и адаптации, тем лучше это выражено. Обрести машущий полёт трудно, для этого должны сойтись многие преадаптации в своеобразных условиях жизни. Зато в дальнейшем поменять рукокрылую специализацию почти невозможно: из летучемышьих крылышек и ножек, носа и уха уже не получишь ни ласты, ни хоботы. В принципе, человек со своей разумностью – из той же серии: возник один раз при странном стечении обстоятельств, достиг своего идеала и не знает, куда эволюционировать дальше.
Рукокрылые, наряду с грызунами, стали одними из главных наших конкурентов, только с другой стороны дерева: если протомыши не пускали приматов на землю и лучше грызли сухие семена, то летучие мыши не дали нам взлететь в воздух и вольготно гоняться там за комарами и бабочками. Осталось прыгать по кронам и искать плоды.
Траво- и листоядные в эоцене в условиях богатейшего развития растительности получили полный карт-бланш.
В Северной Америке и Азии расцвели Dinocerata, или Uintatheria. Твари удались на славу. Монгольские Gobiatherium бултыхались в болотах, рассекая тину огромным носовым гребнем с гигантскими ноздрями. Североамериканские Eobasileus cornutus и Uintatherium anceps (он же Dinoceras), а также китайские Uintatherium insperatus поражают своими метровыми башками: на носу, над глазами и на затылке возвышались огромные костяные шишки-рога, а из пасти торчали длинные изогнутые клыки, которым позавидовал бы любой саблезубый тигр. При этом в целом животные были похожи скорее на длинноногих носорогов и питались растениями. Думается, впечатляющие украшения были созданы половым отбором и использовались для турниров между самцами; саблезубость же типична для очень многих архаичных млекопитающих.
Показательно, что в других линиях копытных отростки на голове чаще возникали не как выросты черепа, а как производные кожи: у оленей они в итоге окостеневают, но площадь их соединения с черепом очень мала, у баранов это роговые образования поверх костяных колышков, у носорогов – утрамбованные волосы. При ударах рог и волосы пружинят, а колебания с них не передаются напрямую на кости мозговой коробки и мозги, а гасятся по пути. Уинтатерии же долбились непосредственно своими черепушками, так что импульс дзынькал прямиком по мозгам. Конечно, наверняка и у них поверх рога была какая-то оболочка, а мозгов было не так уж много, но счастью и процветанию такое устройство черепа явно не способствовало. Из современных копытных максимально к уинтатериевой системе близки жирафы, но они больше бодаются шеями, а не рогами.
На основе афротериев возникло сразу несколько линий крупных травоядных. Родственники эмбритопод Phenacolophidae – Zaisanolophus obscurus в Казахстане и Sanshuilophus zhaoi в Китае – тянутся ниточкой от скромных палеоценовых предков к многообещающим потомкам, а уже ранние Embrithopoda Hypsamasia seni в Турции и Palaeoamasia kansui, Crivadiatherium mackennai и C. iliescui в Румынии представляют развитие группы. Несмотря на принадлежность к афротериям, эта группа скорее продвигалась из Азии в Африку, а не наоборот. Среднеэоценовый намибийский Namatherium blackcrowense с его чрезвычайно широкими скуловыми дугами и высококоронковыми зубами, вероятно, был основателем арсиноитериев, с конца эпохи расселившихся по северной половине Чёрного Континента и достигших успеха уже в олигоцене.
В эоцене появились все главные группы современных афротериев. Достаточно архаично выглядят даманы Hyracoidea, например Seggeurius amourensis и Microhyrax lavocati из Алжира и Марокко и более поздние Bunohyrax matsumotoi из Алжира и Dimaitherium patnaiki из Египта. Видимо, с самого начала они были похожи на сурков. Примитивные даманы были мелкими, их мелкие резцы ещё не росли непрерывно, а теперь они большие и растут, как у грызунов; между резцами и клыками не было зазора-диастемы; клыки ещё иногда были двухкорневые, у современных же этих зубов иногда вовсе нет; лапки были снабжены коготочками, а не копытцами, а ножки были пальцеходящими, а не стопоходящими.
Несмотря на неуклюжий вид и отсутствие хвоста, современные даманы очень здорово лазают по отвесным поверхностям. Для успешного причпокивания к камням и деревьям середины стоп могут подниматься, образуя присоску, а для пущего прилипательного эффекта выделяют резиноподобный пот. Кстати, «я на солнышке лежал, пятки кверху держал, вот они и запотели» – это про даманов, именно так они сбрасывают лишнее тепло, поскольку только подошвы могут потеть. Другая, довольно странная железа расположена на середине спины, она секретирует пахучее вещество при возбуждении; некоторые даманы при опасности оголяют её и поворачивают к хищнику. Показательно, что даманы до сих пор не стали окончательно растительноядными и, кроме травы, листьев и корневищ, регулярно едят насекомых. Рожают даманы от одного до трёх детёнышей, причём очень больших, с самого начала зрячих и пушистых, способных лазать по корягам и камням.
Гораздо более впечатляющие родственники даманов – хоботные Proboscidea. Начав с семнадцатикилограммовых «свинок», они опережающими темпами росли, отъедаясь на сочной болотной растительности Северной Африки. Уже в начале эоцена Daouitherium rebouli вымахал до 80–200 кг, Numidotherium koholense – до 250–300 кг и метра в холке, а на границе с олигоценом – двухметровый Arcanotherium savagei и Barytherium grave – уже до трёх метров в холке! Черепа первых слонов очень похожи на даманьи; головы быстро укорачивались и становились всё более лобастыми; носовое отверстие смещалось на лоб, свидетельствуя о постепенном развитии хобота. Нижние резцы превращались в лопатки на конце всё более узкой нижней челюсти, а верхние – в ещё короткие, но уже узнаваемые бивни.

Moeritherium lyonsi
Наверное, самый известные, хотя и не самые примитивные древние слоны – Moeritherium lyonsi и его родственники из Египта и Ливии. Надо сказать, череп меритерия, если игнорировать нижнюю челюсть и зубы, внешне (с точки зрения антрополога, конечно) довольно похож на череп огромной землеройки – от мелких палеоценовых предков не отвертишься. Зубная формула меритерия была ещё очень примитивной: три резца, клык и по три премоляра и моляра на верхней челюсти, два резца и по три премоляра и моляра на нижней. Короткие толстые бивни, сделанные из вторых резцов, вряд ли торчали изо рта. Сам же зверь был приземистым тапироподобным жителем болот. Слоновья форма мозга меритерия уже вполне узнаваема – кругленькая, укороченная и расширенная, но с огромными обонятельными луковицами.
Файюмский оазис – истинная родина слонов. В нескольких карьерах тут сохранилась богатейшая фауна эоцена и олигоцена – от черепах и крокодилов до китов, слонов и обезьян. Первые находки тут были сделаны Георгом Швейнфуртом в XIX веке, после ещё больше откопал Чарльз Эндрюс, а за ними подтянулись и другие исследователи, которые ныне являются классиками палеонтологии. Файюмские комплексы уникальны наличием и макро-, и микрофауны: чаще сохраняются останки либо больших, либо мелких зверей, а тут есть все. В итоге мы можем прослеживать изменения целой экосистемы на протяжении многих миллионов лет.
Файюм в эоцене и олигоцене представлял собой густо заросшие болота у берега моря, поэтому тут сохранилась и пресноводная, и солоноводная фауна. Удивительно найти посреди Сахары скелеты древнейших китов. Среди наземных млекопитающих огромный процент костей принадлежит именно афротериям, но есть и пришельцы с других континентов – сумчатые, креодонты, антракотерии, грызуны и приматы.
Родственная меритерию Saloumia gorodiskii известна лишь по одному обломку моляра, но важна тем, что жила в Сенегале. Это значит, что наши знания о североафриканских хоботных – лишь частичка реальной мозаики.
На границе эоцена и олигоцена слоны приобрели уже вполне узнаваемый облик: Palaeomastodon beadnelli (Proboscidea) и несколько других видов уже имели хорошо развитые верхние бивни (правда, не слишком длинные и загнутые вниз), утратили лишние резцы и клыки, обрели нормальный хобот и выросли до двух метров в холке. Нижняя челюсть приняла вид длинной лопаты с сошником из соединённых резцов для загребания болотной растительности.
Слоны знамениты самыми разнообразными достоинствами – хоботом, ушами, бивнями, многосменными зубами, огромными мозгами и мощным интеллектом. Но мало кто вспоминает, что у слонов на ручках и ножках есть преполлекс и прехаллюкс – рудименты шестых пальцев (или нулевых, так как они расположены перед первым пальцем). Они развиты, конечно, не совсем полноценно, а в виде сесамоидов (косточек внутри связок), но почти настоящие. Слонам они нужны в качестве задней подпорки под тумбу кисти, когда слон переносит на неё вес тела. Болотная жизнь предков способствовала разлапистости слоновьих кистей и стоп, дабы не увязать в почве; благодаря этому первый палец не исчез, а это дало возможность развития предпервых сесамоидов для дополнительной опоры внутренней стороны кисти и стопы, а это, в свою очередь, обеспечило возможности для ещё большего увеличения размеров тела. У прочих копытных первый палец исчез слишком рано, отчего у них нет таких сесамоидов, что не даёт им стать по-настоящему гигантскими. Впрочем, у некоторых лошадей бывает рудимент большого пальца, так что для них не всё потеряно, надежда на появление многометровых сивок-бурок ещё теплится.
Между прочим, у нас нет никаких доказательств наличия предпервых пальцев у гигантских динозавров зауропод, но у них в силу исходной разлапистой рептильности сохранились другие большие пальцы, так что с опорой у них во все времена было хорошо, что и позволило вымахать до фантастических размеров.
Дополнительные недопальцы встречаются и у других зверей, причём по разным поводам: у панды дополнительный предпервый сесамоид служит для сгребания бамбука; у малой панды имеется такой же, но возникший независимо и конвергентно; у крота предпервый «палец» нужен для копания; у руконожки – для более успешного лазания по ветвям.
Слоны, появившись в болотах, смогли в итоге их покинуть, а вот их родственники сирены Sirenia, напротив, занырнули глубже, уплыв даже в моря. Несмотря на принадлежность к группе афротериев и теоретическую привязку к Африке, древнейшие сирены Prorastomus sirenoides ходили по раннеэоценовым берегам Ямайки. Мордой они были уже вполне узнаваемы, а вытянутые узкие челюсти были снабжены наиполнейшим набором зубов: три резца, клык, пять премоляров и три моляра. В середине эпохи двухметровый Pezosiren portelli на той же Ямайке по-прежнему сохранял хоть и коротенькие, но развитые ноги; тело же было очень длинным, уже вполне дирижабле-сиреновым; совершенно сиреновой была и голова, а зубная формула не поменялась. Хвост пьезосирена не имел увеличенных поперечных отростков, то есть был скорее как у выдры, а не веслообразный. Примитивнейший вид рода Protosiren – P. eothene – обнаруживается в Пакистане в море Тетис, а его многочисленная родня – в Северной Америке, Европе и египетском Файюме: Protosiren fraasi, более крупный и продвинутый P. smithae и другие. Такое универсальное распространение, очевидно, шло вдоль побережий мелководных морей; между прочим, современный дюгонь Dugong dugon тоже живёт вдоль всех берегов Индийского океана – от Меланезии и Австралии до Красного моря и Мадагаскара. Кстати, в том же среднем эоцене примитивные сирены известны и с Мадагаскара – Eotheroides lambondrano, а другие виды этого рода – из Индии и Египта. У испанского Sobrarbesiren cardieli всё ещё были четыре развитые ноги, но задние уже заметно уменьшились. Здорово, что в ряду многочисленных эоценовых и олигоценовых сирен можно проследить все промежуточные стадии уменьшения ног, превращения ручек в ласты, а также изменения морды и зубов.
Современные сирены удивительны множеством особенностей. Они могут жить и в пресной, и в солёной воде и притом, в отличие от всех прочих крупных водных млекопитающих, остаются сугубо растительноядными; оттого у них усложнился желудок и, возможно, поэтому они имеют рекордно маленькое сердце. Задние ноги у них исчезли, хотя рудименты тазовых и бедренных костей всё ещё есть. Передние преобразованы в ласты, в которых, впрочем, вполне сохраняются чёткие сегменты, а у ламантинов намечены пальцы с копытцами. Несмотря на внешнюю редукцию, ламантины своими ручками могут подносить растения ко рту и прижимать детёнышей к груди. Хвост ламантинов имеет вид весла, а у дюгоней раздвоен, как у дельфина. У ламантинов шесть шейных позвонков, в отличие от почти всех млекопитающих, кроме ленивцев. Зубы у ламантинов не имеют внятного числа смен: в каждый момент времени функционируют от пяти до семи зубов, причём на место стёршихся передних сзади появляются новые и сдвигаются, как на конвейере. У дюгоней многочисленные предкоренные и коренные у молодых заменяются на всего пару коренных у взрослых. У ламантинов резцы вырастают, но рано исчезают, замещаясь на роговые пластины; у дюгоней резцы не только сохраняются всю жизнь, но и у самцов превращаются в короткие бивни. У стеллеровой коровы зубы вовсе исчезли, а вслед за ними – и она сама.
Другая линия афротериев пошла по насекомоядному пути: файюмский Dilambdogale gheerbranti стоит в основании тенреков, а Widanelfarasia bowni и W. rasmusseni, хоть и похожи на них же, но представляют, видимо, свою отдельную эволюционную линию. В то же время появляются и древнейшие прыгунчики, например Chambius kasserinensis, Nementchatherium senarhense и Herodotius pattersoni.
Современные тенреки Tenrecoidea, или Tenrecomorpha, – очень своеобразные зверюшки. Большая их часть живёт на Мадагаскаре, но некоторые – в материковой Африке. Многие мадагаскарские тенреки конвергентны ежам: обыкновенный тенрек Tenrec ecaudatus выглядит как бесхвостый опоссум со спрятанными в шерсти иголками, полосатый тенрек Hemicentetes semispinosus – как чёрно-жёлтый недоёжик с мордочкой-трубочкой, редкими иголками и растрёпанным колючим хохлом на загривке, а ежиный тенрек Setifer setosus и малый тенрек Echinops telfari вообще внешне от ежей малоотличимы. Живут тенреки как на земле, в том числе в норах, так и на деревьях, питаются в основном насекомыми, но при случае не отказываются и от фруктов. Тенреки способны впадать в спячку в сухой сезон. Тенреки – рекордсмены среди плацентарных по плодовитости, самки могут иметь до 24 сосков и родить до 25 детёнышей! Это, кроме прочего, стало одним из тормозов на пути поумнения тенреков: на каждого детёныша приходится слишком уж мало мозга и родительского внимания, социальность у тенреков совсем слаба и ограничивается сбиванием в кучку в каком-нибудь дупле для согревания ночью.
Среди тенреков есть и масса других экологических вариантов: длиннохвостые тенреки Microgale больше всего похожи на землероек с чрезвычайно длинным хвостом, рисовые тенреки Oryzorictes – на помесь жирной землеройки с длиннохвостым кротом, болотные тенреки Limnogale mergulus – на водяную крысу с лысым хвостом и перепонками на задних лапках, они хорошо плавают и питаются водными растениями и рыбой. Выдровые землеройки Potamogale velox, Micropotamogale lamottei и M. ruwenzorii в Экваториальной Африке внешне и по образу жизни скорее похожи на выдр и питаются соответственно – раками, рыбой и лягушками.
Очевидно, мадагаскарская экосистема, столь бедная на зверей, взрастила спектр экологических вариантов из того, что было в распоряжении. Заодно, глядя на разнообразие тенреков, можно легко представить раннекайнозойскую эволюционную радиацию млекопитающих.
Прыгунчики Macroscelidea частично конвергентны тушканчикам: у них длинные тонюсенькие ножки, длинный тонюсенький хвостик и компактное пушистое тельце. Нос прыгунчиков превращён в смешной вечно-подвижный хоботок. А вот зубы их мало отличаются от зубов насекомоядных, в которых прыгунчики традиционно и включались. Образцом прыгунчиков может служить короткоухий прыгучик Macroscelides proboscideus, а чёрно-оранжевые хоботковые собачки Rhynchocyon даже очень красивы. По некоторым чертам строения зубов и черепа прыгунчики похожи на тупай, а иногда могут болеть малярией, что тоже отсылает к приматам. Между прочим, головной мозг прыгунчиков развит очень даже хорошо относительно размера тела и челюстей; прыгунчики очень бодры и постоянно находятся в движении, у них отлично развиты все органы чувств. В принципе, из такого первоисходника можно было бы получить кого-то и более разумного, тем более что и самка рождает обычно всего одного-двух детёнышей. Только вот специализация к прыганию и рытью подвела: хватательную и трудовую конечность из прыгунчиковой лапки не сделаешь! К тому же детёныши прыгунчиков рождаются практически готовыми – пушистыми, зрячими и способными бегать. Всё детство занимает пару месяцев, так что на обучение времени совсем нет; продолжительность же жизни, как, кстати, и у тенреков, всего год-полтора.
В то время как афротерии превращались в тварей от прыгунчиков до слонов, прочие плацентарные – бореэвтерии – эволюционировали по-своему. На протяжении всего эоцена продолжали бытовать поздние кондиляртры Condylarthra типа Hyopsodus и Phenacodus, похожие на помесь куницы с карликовым оленьком. Некоторые из них ещё могли лазать по деревьям, некоторые рыли норы, но всё больше и больше переходили к бегу по кустам и растительноядности. Показательно примитивное строение их мозга: новая кора была слабо распространена вниз и не имела борозд с извилинами, обонятельные луковицы были огромными, а мозжечок – маленьким.
Удивительно обнаружение Tingamarra porterorum – кондиляртры из раннего эоцена Австралии. Пусть это лишь один-единственный моляр, но чрезвычайно важный, так как показывает возможность миграции плацентарных зверей из Южной Америки в Австралию. Чисто гипотетически Австралия могла быть заселена кондиляртрами, из которых могли бы возникнуть самые разнообразные оригинальные существа. Однако некоторые палеонтологи считают отложения с зубом тингамарры позднеэоценовыми или даже позднеолигоценовыми, а зуб – принадлежащим сумчатому типа примитивного опоссума. Тогда вся красивая гипотеза осыпается. Возможно, новые находки в будущем лучше прояснят загадку тингамарры.
Южноамериканские копытные эволюционировали своими путями. Особенно странно выглядела аргентинская Thomashuxleya externa из группы Notoungulata, одна из первых представительниц Toxodontia. Это было животное весом 180 кг с мордой клыкастой собаки-дегенератки, на крепких лапах, с маленьким хвостиком. Судя по зубам и челюстям, питалась она, видимо, растениями, хотя по внешнему виду это было неочевидно. Другие нотоунгуляты были странны по-своему: у Notostylops зубная формула могла меняться от 3143/3143 до 2033/2033. Описано множество видов нотостилопсов, но есть мнение, что дело не в систематике, а в величайшей индивидуальной изменчивости. В целом эти зверюшки были очень усреднёнными, генерализованными: собакоподобные твари размером с кролика, экологически что-то вроде всеядных енотов, но с резцами, как у грызунов.
Зубная формула отражает число зубов в зубном ряду верхней и, через дробь, нижней челюсти; зубы считаются только на одной стороне. Первой цифрой идут резцы, потом клыки, затем премоляры и последними – моляры.
Нотоунгуляты Pseudhyrax eutrachytheroides и P. strangulatus были аналогами даманов.
Маленький нотоунгулят Notopithecus adapinus первоначально был описан как примат, однако в реальности был копытным, похожим на длинноногую мордастую крысу, неплохо лазавшую по деревьям.
В эоцене Аргентины появились также первые представители отряда Litopterna (или Notopterna) – Amilnedwardsia brevicula и Indalecia grandensis, родственные знаменитым и гораздо более поздним макраухениям.
Доживали свой век и последние представители Xenungulata – Etayoa bacatensis в Колумбии размером с собаку и более крупная Notoetayoa gargantuai из Аргентины.
Кроме Австралии, Африки и Южной Америки существовал и ещё один материк-остров – Индия, плававшая в эоцене посреди моря Тетис. Тут развились свои аналоги хоботных и американских копытных – отряд Anthracobunia, например, многочисленные виды Anthracobune и Cambaytherium. Антракобуниды настолько похожи на первых хоботных, что некоторые авторы считают их предками слонов. Другие, впрочем, определяют их лишь как семейство Anthracobunidae в пределах непарнокопытных. Как и большинство эоценовых растительноядных, антракобуниды внешне больше всего напоминали тапиров, но имели довольно стройные ноги и длинный тонкий хвост. Интересно, что, несмотря на изоляцию, многие другие животные эоценовых фаун Индии родственны европейским, африканским и, в меньшей степени, азиатским. Однако антракобуниды – эндемики Индии, они так и не смогли переселиться куда-то ещё и в последующем вымерли, видимо, не выдержав конкуренции с привычными нам копытными.
Североамериканские и евразийские копытные пошли двумя основными путями – непарно- и парнокопытным.
Различаются непарно- и парнокопытные, конечно, не только парностью пальцев на ножках, хотя этот признак был важен и надёжен уже в эоцене. Две эти группы сделали разные ставки на развитие жевательного аппарата и пищеварительного тракта: непарнокопытные стали гораздо эффективнее жевать первично и переваривать кишечником, а парнокопытные – жевать вторично и переваривать желудком. Проще говоря: лошадка сначала старательно жуёт, потом немножко переваривает желудком, а после долго – кишечником, а коровка – сначала глотает не жуя, потом частично переваривает желудком, потом отрыгивает и тщательно пережёвывает, потом снова глотает и снова переваривает желудком, а потом ещё и кишечником. В итоге у непарнокопытных однокамерный желудок и очень длинный и важный кишечник, а у парнокопытных – четырёхкамерный желудок и сравнительно менее значимый кишечник.
Эоценовые непарнокопытные быстро взяли курс на увеличение размеров и были склонны давать гигантские формы, тогда как парнокопытные чаще оставались мелочью.
Непарнокопытные Perissodactyla начали свой подъём к успеху. Первые представители – Palaeotheriidae в лице английского Hyracotherium leporinum, немецкого Propalaeotherium parvulum и французского Palaeotherium magnum, что логично, мало отличались от кондиляртр: размером с большую кошку или собаку обитатели густых зарослей, питающиеся листьями кустарников и плодами. На передних ногах у них было четыре пальца, а на задних – уже всего три; на низких молярах – шесть тупых бугорков без всяких режущих гребней. В дальнейшем они пошли несколькими альтернативными путями.
Иногда из палеотериев выделяется семейство Pachynolophidae, представленное, например, Pachynolophus и Anchilophus. Эти звери были доминирующими непарнокопытными среднего эоцена Европы, но к концу эпохи очень резко сократились в числе и к началу олигоцена полностью вымерли.
Многие ранние непарнокопытные по славной эоценовой традиции стали тапироподобными жителями болот. Группа тапиров в виде нескольких архаичных семейств возникла в Северной Америке. Это и древнейшие мелкие Heptodon, и стройные Helaletes nanus. Довольно быстро они попали в Азию, где известны от Монголии до Казахстана, например Desmatotherium. Каким-то образом они добрались и до острова Индии – Vastanolophus holbrooki и Cambaylophus vastanensis. Первобытные тапиры обычно были довольно мелкими и ещё не имели такого хобота, как современные, но принципиально схожи со своими потомками. Между прочим, несмотря на название отряда, тапиры по-прежнему имеют четыре пальца на передних ногах, но три – на задних.
Особняком стоят азиатские эоценовые тапиры Lophialetes expeditus и Schlosseria magister, имевшие по три пальца и на стопе, и на кисти. Лофиалетесы вообще обогнали своё время, благо обгонять они умели: в эпоху господства толстеньких пожирателей околоводной растительности лофиалетесы были быстробегающими стройными газелеподобными длинношеими потребителями высоко расположенных листьев открывающихся степей. Однако, несмотря на прогрессивность, лофиалетесы вымерли, не выдержав конкуренции с парнокопытными, а более архаичные формы тапиров сохранились.
Тапироподобны были и первые носороги – Yimengia, Triplopus, Hyrachyus, Lophiodon, Amynodon, Metamynodon и прочие, относящиеся к нескольким архаичным семействам. Многие из этих родов имели крайне широкое распространение – в Северной Америке, Европе и Азии. Amynodon известен также из Японии, а Hyrachyus – даже на Ямайке! Большинство из них были приземистыми бочкообразными коротконогими жителями болот и прибрежных зарослей и питались ветками и листьями кустарников. В отличие от тапиров, носороги сделали ставку на режущие свойства зубов, высококоронковость, большие размеры тела и, как это ни противоречит предыдущему пункту, – на способности к бегу. У некоторых архаичных носорогов сохранялись нормальные резцы и довольно большие клыки; на ногах часто по-прежнему было четыре пальца спереди и три – сзади; степные лошадеподобные Hyracodon и предки индрикотериев Forstercooperia уже сократили число пальцев на кисти до трёх. Между прочим, китайские среднеэоценовые индрикотерии Juxia приобрели свой классический облик, только размером были скромнее своих знаменитых олигоценовых потомков. Судя по отступанию носовых костей назад, у продвинутых Cadurcodon имелся небольшой хобот; эти болотные животные максимально походили на тапиров, да к тому же с большими вертикальными бивнями. Вот чего не было у первых носорогов, так это рогов! Не было их даже у первых настоящих носорогов семейства Rhinocerotidae – среднеэоценовых североамериканских Uintaceras radinscyi.
Вот у кого рога развились, так это у Brontotheriidae, большая часть которых известна из Северной Америки; между прочим, филогенетически бронтотерии родственнее лошадям, а не носорогам. Самые первые мелкие Eotitanops ещё имели простенькую морду, чуть более поздние Palaeosyops размером с корову и ещё более мощные Manteoceras уже обзавелись вздутием на носу, а Protitanops curryi, Protitan имели хотя бы и невысокие, зато раздвоенные рога на носу. Бронтотерии, начав, как и все, с размера карликовой свинки, быстро выросли в огромных чудищ до двух с половиной метров в холке, став самыми большими и массивными животными своего времени. Огромные остистые отростки первых грудных позвонков были нужны для крепления мощнейшей мускулатуры для поддержания тяжеленной головы, тому же соответствовали и толстые ноги. В итоге вид бронтотериев был куда как харизматичный. Самый широко-разрекламированный бронтотерий – Megacerops (он же Brontotherium и Brontops), хотя пара его тупых округлых рогов на оригинальных черепах выглядит далеко не так впечатляюще, как на реконструкциях. У самцов рога были больше, чем у самок, что уже намекает на половой отбор и брачные турниры. Как и в случае с уинтатериями, бодание костными рогами должно было создавать сложности для сохранности мозга; впрочем, миллионы лет бронтотериям это проблемой не казалось. Известен скелет бронтотерия с результатом неудачного брачного турнира – сломанным и заросшим с костяной мозолью ребром на правом боку.

Megacerops
Монгольские Embolotherium пошли куда дальше: их носовой рог стал широченной раздвоенной лопатой, причём одинаково большой и у самцов, и у самок. Борозда на передней стороне шла от носовой полости и, возможно, служила для размещения резонатора: монгольские болота оглашались трубными кликами эмболотериев. Судя по румынскому Brachydiastematherium transylvanicum, подобные монстры жили и в Европе.
Эволюция лошадей – классика палеонтологии, каждый видел в школьном учебнике схему их эволюции, составленную В. О. Ковалевским. А начинается она с североамериканского Eohippus angustidens – первого представителя семейства Equidae. Эогиппус очень мало отличался от гиракотерия (а иногда они даже объединяются): всего 30 см в холке, со среднюю собачонку. Коронки его зубов были низкими, на ногах, как у всех примитивных непарнокопытных, по четыре пальца на передних и по три – на задних, из чего следует, что жил он в густых зарослях, бегал по влажной почве и питался листвой и тонкими ветками.

Eohippus angustidens
По мере изменения условий жизни исходная «кондиляртность» уменьшалась, а «лошадность» усиливалась, причём в нескольких эволюционных линиях независимо и немножко по-разному. Лошадиная ветвь представлена, например, последовательными Orohippus и Epihippus. У них всё ещё имелись по четыре пальца спереди и три сзади, а локтевая и лучевая кости предплечья оставались независимыми в память о роющих предках; по-прежнему низкокоронковые зубы помаленьку обзаводились гребнями для разрезания листьев; размеры тела помаленьку росли.
В. О. Ковалевский на основе тщательнейшего изучения эволюции конечностей и зубов ископаемых лошадей и энтелодонтов пришёл к выводу, что, во-первых, разные части тела эволюционируют с неравными скоростями и, во-вторых, в случае обгона одной системы другой общая итоговая приспособленность может оказаться ниже, чем у конкурентов, у которых признаки менялись медленнее, но равномернее. В. О. Ковалевский разделил понятия инадаптивной эволюции – неравномерной и не вполне согласованной, накладывающей ограничения на последующие прогрессивные изменения, и адаптивной – более соразмерной, без крайностей и заскоков. Кроме того, В. О. Ковалевский первым в палеонтологии развил и последовательно применял морфофункциональный анализ – оценку эволюционных изменений с позиции их приспособительного смысла в данной конкретной экологической среде. Сейчас такой подход кажется само собой разумеющимся, но в XIX веке это был мощнейший прорыв; стоит ведь помнить, что эволюционная теория только-только была опубликована, а палеонтология возникала на глазах. Да и сейчас во многих палеонтологических трудах занудное перечисление морфологических отличий следует без малейшей попытки осознать их смысл.
В эоцене среди лошадеобразных выделились Eomoropidae, например Paleomoropus jepseni и Eomoropus из Восточной Азии и Северной Америки. Эти листоядные звери размером с овцу имели довольно крупные клыки в пасти и когти на пальцах и могут быть предками более поздних халикотериев. Некоторые подобные животные, например европейский Lophiaspis, иногда уже напрямую относятся к халикотериям; впрочем, эоморопиды могли быть и параллельно развивавшейся группой.
Примитивные парнокопытные Artiodactyla в раннем эоцене эволюционировали несколько медленнее, чем непарнокопытные, зато раскачались к концу эпохи. Первые представители – Dichobunidae в лице многих видов раннеэоценовых Diacodexis, среднеэоценовых Homacodon и прочих других – широчайше распространились по Европе, Северной Америке и Азии. Внешне они выглядели как карликовые оленьки с длинными тонкими хвостиками весом два-три килограмма; несмотря на принадлежность к парнокопытным, они имели ключицы и по пять пальцев с копытцами на каждой ножке. Подобны им, разве что крупнее и с короткими хвостиками, были и североамериканские Hypertragulidae. Удивительна рассинхронизированность эволюции передних и задних конечностей: у Hypertragulus всё ещё пятипалая когтистая кисть сочеталась с функционально двупалой стопой (сохранялись рудиментарные фаланги ещё одного пальца стопы, но совсем крошечные), а у Leptomeryx – четырёхпалая ручка с полностью двупалой ножкой. Показательны изменения зубов: верхние резцы уже успели исчезнуть, но у гипертрагулюса имелись довольно крупные клыки и острые премоляры, а нижние резцы были почти вертикальны, тогда как у лептомерикса клыки уменьшились, премоляры почти перестали отличаться от моляров, а нижние резцы направились горизонтально вперёд. Несмотря на примитивность, гипертрагулюсы дожили до миоцена.
Азиатские Archaeomeryx optatus и его родственники размером с зайца были и внешне похожи на зайцев: попрыгунчики с длинными задними и короткими передними ножками-шпильками, развитыми верхними резцами, но ещё и с клыками, и с длинным хвостом. Очень быстро, уже к концу эоцена, подобные существа доэволюционировали до семейства Tragulidae, примерами чего могут быть Archaeotragulus krabiensis и Krabitherium waileki из Таиланда.
Современные карликовые оленьки Tragulidae – азиатские Tragulus и Moschiola и африканские Hyemoschus aquaticus – просто напрашиваются на то, чтобы стать объектом загадок: саблезубый зверь, отлично плавает и даже ходит по дну, где ловит рыбу и раков, лазает по деревьям и лианам, день проводит в дупле, ночами выходит на поиски листьев, плодов, крыс, ящериц и всяческой падали. Кроме огромных клыков у самцов, оленьки обладают невероятно длинным языком и трёхкамерным желудком, а никаких намёков на рога у них нет. Весят они от полкило до восьми килограмм, африканские гиганты – до пятнадцати. По итогу, оленьки – едва ли не самые примитивные из нынешних парнокопытных, в их лице можно видеть осколок эоценового мира влажных тропических джунглей.
Саблезубость присуща и некоторым другим парнокопытным, например кабарге Moschus moschiferus, водяному оленю Hydropotes inermis и мунтжаку Muntiacus muntjak, причём у последнего клыки сочетаются с рогами.
Успешной и многочисленной группой эоценовых парнокопытных были Anthracotheriidae, например, древнейший мелкий Elomeryx и поздний двухсоткилограммовый Bothriodon (он же Ancodus); появились они, видимо, в Азии, потом оказались в Европе, а оттуда добрались и до Северной Америки в одну сторону и Африки – в другую. Особенно преуспели многочисленные виды Anthracotherium, дожившие от среднего эоцена аж до миоцена. Эти звери выглядели как приземистые тонконогие свино-тапиро-бегемотики с вытянутыми мордами, и ведь вправду они оказались предками гиппопотамов, а по менее вероятной версии – родственниками свиней. Набор зубов был практически полным, на передних лапках имелось по четыре пальца, а на задних – по пять. Такое строение оказалось практически универсальным для жизни в заросших болотах. Впрочем, антракотерии ещё не были слишком специализированы и могли уклоняться в разные стороны: некрупный бирманский Siamotherium pondaungensis бегал по открытым лесам и был вполне всеядным, не слишком уважая листья. Видимо, подобные антракотерии или очень родственные животные стали основой и для монстрических энтелодонов, и, возможно, для китов.

Anthracotherium
Современные копытные рождаются практически готовыми, работоспособными, иногда они даже почти сразу пытаются щипать траву, хотя, конечно, довольно долго ещё кормятся молоком. Повышенная самостоятельность резко повышает выживаемость, даже при небольшом числе детёнышей – у копытных обычно один на одни роды. Получается, что, с одной стороны, это высшая степень доделанности и прогресса на момент рождения среди млекопитающих, а с другой – парадоксальным образом копытные вернулись в идеал рептилий, у которых из яйца вылупляется уже готовая ящерка, только маленькая. По сравнению с примитивными млекопитающими, то есть нами, такая стратегия вроде как и продвинутая, но диалектически становится тормозом на пути развития разума, ведь если детёныш обеспечен с рождения всеми необходимыми программами поведения, то и обучение уже не нужно. Оттого мы – архаичны по сравнению с овцами, но мы – люди, а они – овцы.
Во второй половине эоцена часть парнокопытных Северной Америки стала увеличиваться в размере, превратившись в двухметровых кабаноподобных тварей Achaenodon – самых крупных парнокопытных в своих фаунах. Они были либо первыми и уже впечатляющими Entelodontidae, либо, вероятнее, гигантскими представителями Helohyidae. Ахенодоны сохранили весьма архаичную зубную систему, в том числе огромные толстые клыки, острые премоляры и бунодонтные – низкокоронковые бугорчатые – моляры, а их короткие массивные головы были чем-то средним между кондиляртровыми и свиными. Вероятно, эти монстры были не сугубо растительноядными, а хотя бы частично занимали нишу крупных падальщиков и могли грызть кости. Несколько позже их сместили с пьедестала Brachyhyops и Archaeotherium – уже несомненные представители энтелодонтов, расселившиеся и по Северной Америке, и по Азии, и по Европе, успешно перешедшие в олигоцен и давшие там великолепное потомство. Длинные челюсти археотериев были усажены примитивными зубами, в том числе огромными клыками, и снабжены крупными выростами для мощнейших жевательных мышц, о чём явно свидетельствуют и широкие скуловые дуги. Как и ахенодоны, археотерии, кроме растений, пожирали падаль; этому же способствовало и отличное обоняние, о чём мы можем судить по увеличенным обонятельным луковицам. Тяжеленная голова поддерживалась сильными мышцами, крепящимися на длинные остистые отростки грудных позвонков, отчего археотерии имели ссутуленный вид. Ноги, несмотря на сравнительно большую длину, не были приспособлены к быстрому бегу, а кисть и стопа адаптировались к ходьбе по влажному грунту. Эти твари стали конкурентами не только и столько растительноядным, сколько мезонихиям и креодонтам.
Отдельная ветвь парнокопытных – мозоленогие Tylopoda. Они настолько специфичны, что иногда выделяются в самостоятельный отряд. Называть их «парнокопытными» даже язык не поворачивается, ведь на пальцах у них не копыта, а толстые кривые когти, а кисть и стопа у современных – пальцеходящие, а не фалангоходящие; имеются у них и другие уникальные особенности, например овальные эритроциты, специфика зародышевых оболочек и строения желудка. Современные мозоленогие довольно однообразны – азиатские верблюды и южноамериканские ламы, а вот с эоцена по миоцен эта группа была многообещающа. Совсем не похожи на верблюдов низкорослые короткомордые Oreodonta (иногда определяются как Oreodontoidea, или Merycoidodontoidea), например североамериканские Protoreodon, Agriochoerus и Merycoidodon. По внешности это было что-то среднее между полутораметровой свиньёй и бегемотом, но с почти верблюжьими зубами и длинным тонким хвостом. Верхние клыки сочетались с клыкоподобным первым нижним премоляром, тогда как нижний клык был резцеподобным. Судя по специфической ямке на черепе впереди от глазницы, ореодонты могли иметь пахучие железы для мечения территории; о том же косвенно свидетельствует отличное развитие обонятельных частей мозга, например у Bathygenys reevesi. На кистях и стопах было по четыре длинных функциональных пальца и один маленький рудиментарный, все с когтями или копытцами; кости предплечья и голени не срастались. Несмотря на бочкообразность тела, коротконогость и общую примитивность конечностей, ореодонты были одними из первых жителей степей, где паслись их стада, состоящие из сотен животных, что видно по количеству находок. По факту ореодонты были едва ли не самыми массовыми животными своего времени. Надо думать, в последующем это благолепие истребили новые хищники и вытеснили более шустрые копытные.

Merycoidodon
Другие североамериканские мозоленогие – Oromerycidae, например Protylopus размером с зайца и Eotylopus величиной со среднюю антилопу, – сохраняли самый генерализованный облик и примитивнейшую зубную систему, зато в их предплечье локтевая срослась с лучевой, а в голени малая берцовая редуцировалась до невнятной шпильки. Эоценовые североамериканские Protoceratidae – мелкие Leptotragulus и Leptoreodon, а также более крупные Heteromeryx dispar и Pseudoprotoceras – были предками самых необычных рогатых верблюдов более поздних эпох; пока же они не слишком выделялись на общем фоне. Наконец, в эоцене появляются и настоящие верблюды Camelidae – североамериканские Poebrodon, известные лишь по нескольким зубам; будущее этой группы было впереди.
Совсем другим путём пошли родственники ореодонтов и верблюдов в Европе. Маленькие низенькие коротко-тонконогие и очень длиннохвостые Cebochoeridae, например Gervachoerus, сохраняли генерализованный кондиляртровый облик. У Choeropotamidae – Amphirhagatherium – хвостик укоротился, а задние ножки стали сильно длиннее передних, обеспечивая попрыгучесть; судя по бунодонтным зубам, они питались фруктами. Дальше всех пошли Cainotheriidae, например Cainotherium: от эоцена до миоцена эти крошечные – 15 см в высоту – копытные «зайчики» прыгали по зарослям и открытым степям. Несмотря на подвижность, кайнотерии сохраняли по четыре пальца на ручках и ножках. Кроликоподобным стало не только телосложение, но и череп, и зубы.
Серьёзнее выглядели европейские Anoplotheriidae, например Anoplotherium – очередные тапироподобные почти-мозоленогие с пятью толстыми пальцами на передних и четырьмя на задних коротких ножках, с длинной мордой и толстым кенгуруподобным хвостом. Предполагается, что аноплотерии могли вставать на задние ноги, вытягиваясь за высокорастущими листьями. Xiphodontidae в лице Xiphodon и его родни представлены, наоборот, длинноногими стройными двупалыми подобиями лам.
Самые нестандартные парнокопытные среднего эоцена с самой нестандартной судьбой – Raoellidae в лице индийских Indohyus indirae и I. major. Эти мелкие зверюшки на тонких ножках оказываются ближайшими родственниками китов! Как ни странно, это вполне можно видеть на тварюшке меньше метра длиной. Но половину этой длины составляет мощный хвост, тонюсенькие косточки имеют чрезвычайно толстые стенки, а крошечная черепушка выглядит как арифметическое среднее между свиной и дельфиньей. Особенно же специфично утолщение медиальной стенки тимпанической пластинки височной кости при уменьшении толщины латеральной стенки: такая форма встречается только у китообразных, и случайное совпадение по столь особенному признаку маловероятно. Эти и другие особенности свидетельствуют об отличных способностях к нырянию; возможно, в воде индохиусы спасались от хищников, как это делают африканские карликовые оленьки, способные задерживать дыхание на четыре минуты. Судя по зубам, индохиусы были растительноядными или всеядными и точно отличались в этом отношении от самых первых китов. Более того, индохиусы явно не могли быть прямыми предками китов, ведь древнейшие китообразные уже существовали и даже найдены в тех же местонахождениях, но на примере этих тварюшек мы воочию видим «живых ископаемых» переходных форм.

Indohyus indirae
Вообще, эволюция китообразных Cetacea – удивительнейшая тема. Начать с того, что Гималаи – родина китов. Казалось бы, как могли величайшие вершины мира породить самых водных зверей? Но в эоцене гор ещё не было, а их будущие заоблачные хребты были дном моря Тетис. А на берегах и мелководьях жили будущие киты. Кто конкретно из великого изобилия древнейших копытных был прямым предком – до сих пор вопрос. Генетические данные указывают на бегемотов как ближайших современных родственников, но бегемоты – это выжившая родня, они появились гораздо позже, да к тому же в Африке.
Одни из лучших кандидатов на роль предков китов – копытные хищники Mesonychia. В эоцене они расцвели пышным цветом, дав крайне харизматичных представителей: Mesonyx с приличного волка и такого же облика; масса видов гиеноподобных, но размером с медведя, Harpagolestes с широчайшим распространением от Северной Америки до Центральной Азии; Pachyaena, среди нескольких видов которой рекордных значений достигла P. gigantea – под сотню килограмм веса, а по смелым оценкам – даже до четырёхсот! В отличие от всех прочих хищников эоцена, мезонихии, будучи копытными, здорово умели бегать и, несмотря на традиционно тапироподобное тело, были хотя и не быстрыми, зато выносливыми. Также, мезонихии не развили специализированных хищнических зубов, их коренные зубы тупы, так что они могли быть не столько хищниками, сколько падальщиками.
Совсем особняком стоит Andrewsarchus mongoliensis из Монголии – крупнейший хищный зверь всех времён, со скуластым черепом 83 см длиной и 56 см шириной! На всякий случай, у белого медведя максимальная длина черепа 44 см, а у пещерного – 49 см. На таком фоне даже огромные клыки эндрюсархуса не кажутся огромными; а ведь клыкоподобным был ещё и второй резец, отделённый от обычного клыка маленьким третьим резцом. Одновременно моляры эндрюсархуса совсем тупые. Сочетание длинного зубного ряда без зазоров, широкой глотки и способности крайне широко открывать пасть говорит о всеядности зверя. Растопыренные вверх и в стороны глазки косвенно свидетельствуют об околоводной жизни. Проблема в том, что от эндрюсархуса сохранился лишь один-единственный череп, так что его истинный облик и образ жизни остаются под вопросом. Таксономическое положение эндрюсархуса загадочно, он родственен то ли мезонихиям, то ли энтелодонтам, то ли китам. Длинный приплюснутый крокодилообразный череп весьма напоминает черепа самых первых китов, так что эндрюсархус мог быть своеобразным сухопутным китом.
Другое дело, что в некоторый момент генетика показала родство китообразных и современных бегемотов, – и с этого момента многие специалисты резко поменяли своё мнение. Генетика – это же не скучная анатомия, гены не видать, стало быть, их анализ – это высшая степень магии, а потому даже многие прожжённые морфологи склонны доверять генетическим данным больше, чем собственным глазам. Крайне показательно, что в кладограммах, построенных на основе морфологии, мезонихии чаще оказываются ближайшей роднёй китов, но при включении в анализ молекулярных данных на первый план выходят бегемоты, а мезонихии удаляются в родню креодонтов и хищных. Но постойте! Ведь у нас нет вообще никаких сведений о генах мезонихий! Так может, дело не в родстве, а в тонкостях статистических методов и допущениях с предположениями? К тому же, строго говоря, одно не так уж исключает другое: черепа антракотериев, бегемотов, мезонихий и китов все так или иначе напоминают друг друга, так как восходят к кондиляртровому исходнику – низкие, плоские, с длинными вытянутыми челюстями, суженными спереди и широкими сзади, с огромными развёрнутыми скуловыми дугами, с тенденцией к увеличению клыков, с глазками, расположенными сбоку-сверху. И от такого неспецифического кондиляртрового корня легко выйти и на хищных мезонихий, и на растительноядных индохиусов с антракотериями и бегемотами, и на рыбоядных китов. В разных линиях сохранялись и усиливались разные черты, да к тому же добавлялись новые.
Есть несколько версий, как и зачем киты вообще полезли в воду.
Если принять травоядную версию предков, то мелкие оленькоподобные зверьки могли просто прятаться под водой от злых хищников, а потом увлеклись нырянием. При мелких размерах, исходной клыкастости и недалёкости от насекомоядных исходников у них уже была некоторая склонность к мясоедству, примером чему служат современные карликовые оленьки, ловящие мелких животных, причём часто именно рыбу. И вот сегодня ты, трепеща, захлёбываешься на дне речки, спасаясь от кровожадного креодонта, рыщущего по берегу, а завтра сам с удовольствием лопаешь селёдку в океане.
Если же предками всё же были мезонихии, то они уже были хищниками и падальщиками. Такие твари могли собирать всякие вкусняшки на берегах водоёмов, а потом увлечься серебристой чешуёй – и уплыть за горизонт.
Если первые этапы окитения пока ещё скрываются в палеонтологическом тумане, последовавшая эволюция изучена просто отлично: известны все детали преобразования всех частей тела. Радостно наблюдать, как от одного вида к другому зверюшки меняются: стартовав 53–48 млн л. н., четвероногие собакоподобные ко времени 48–47 млн л. н. становятся всё более ластолапыми и тюленеподобными, к 42–37 млн л. н. – хорошо китообразными, но всё ещё с ножками, а около 34 млн л. н. – полноценными китами современного облика. Таким образом, на полное преображение понадобилось от десяти до двадцати миллионов лет – срок, вполне сопоставимый с появлением черепах из ящерицеподобных предков и прямоходящих разумных людей из совсем-совсем обезьян. Видимо, такой темп эволюции – более-менее норма, определяемая терпимым давлением отбора и скоростью появления удачных мутаций.
Древнейшие – ранне-среднеэоценовые – достоверные киты выделяются в семейство Pakicetidae; все они найдены на границе Пакистана и Индии. Сейчас их известно уже не так мало: Ichthyolestes pinfoldi размером с лису, Pakicetus attocki, P. inachus и другие виды – с собаку, Nalacetus ratimitus – с волка. У всех них отлично развиты и даже длинны все четыре ноги, кисть даже пятипалая, стопа – четырёхпалая. У них сохранялись ключица и третий вертел бедренной кости, а ведь они имелись лишь у самых ранних и примитивных парнокопытных. Шея была не то чтобы длинной, но нормальной. В черепе поражает мизерная заглазничная ширина, так что мозговая коробка соединялась с челюстями тоненьким стебельком, при огромных широко развёрнутых скуловых дугах. Вытянутые челюсти оборудованы гетеродонтными зубами, причём заклыковые зубы имеют выраженные бугорки; ноздри открывались на кончике морды. Почему же их относят к китам? Очень толстые кости приспособлены к нырянию; очень длинный и массивный хвост – залог будущего китовьего хвоста с лопастями; глаза расположены сбоку и сверху – опять же для ныряния; в слуховой капсуле медиальная стенка утолщена, как у китов, хотя полукружные каналы большие (у более поздних китов они малы), да и в остальном внутреннее ухо устроено скорее как у наземных животных, так что пакицеты неплохо слышали и в воде, и на суше. В итоге почти все особенности пакицетов промежуточны между наземным копытно-мезонихиевым состоянием и водно-китовьим. Судя по зубам, пакицеты уже могли быть рыбоядными, хотя по тем же зубам видно, что предки явно были растительно- и всеядными. Любопытно, что налацет найден в пресноводных речных отложениях, а не в морских, так что приспособление к нырянию и плаванию началось ещё до освоения морских побережий.
Гораздо более водным был раннеэоценовый пакистанский Ambulocetus natans, с парой родственников выделяемый в семейство Ambulocetidae (индийский Himalayacetus subathuensis древнее даже пакицетов, но известен лишь по фрагментам челюстей). Размер вырос вдвое – до трёх метров. В черепе амбулоцетуса китообразность уже вполне узнаётся, редко стоящие зубы по-прежнему делились на классы, премоляры сохраняли два корня, а моляры заострились, но всё ещё были довольно широкими. Передние лапы укоротились, хотя сохранили пять пальцев, а задние ноги оставались довольно длинными. На таких лапках по суше можно было передвигаться наподобие сивучей и моржей.
С этого момента стали возникать варианты, например индийские Remingtononocetus domandaensis и R. harudiensis отличаются чрезвычайно длинной низкой и узкой головой, подобной гавиальей. Тут уже в рыбоядной специализации сомнений быть не может.
Лучше всего удалась среднеэоценовая группа Protocetidae, сейчас их известно уже полтора десятка родов. Трёхметровый Maiacetus inuus замечателен не только своей примитивностью, но и тем, что в скелете самки сохранился скелет детёныша. Он ориентирован так, что роды должны были проходить головой вперёд, как у наземных млекопитающих и в отличие от китов, у которых детёныши рождаются вперёд хвостом. Впрочем, есть сомнения – а не съела ли «кит-мать» этого несчастного детёныша? Следов зубов на костях вроде и нет, но расположение маленького скелета в грудной клетке большого как-то не вполне соответствует расположению матки. Благодаря отличной сохранности майяцетуса и других родственных китов – Artiocetus clavis, Rodhocetus kasrani, R. balochistanensis и прочих, мы знаем, что их кисти и стопы были очень похожи на кисти и стопы примитивных копытных. Череп не принципиально изменился по сравнению с предками, протоцетусы по-прежнему имели неплохое обоняние, но уже хорошо слышали под водой; задние зубы всё ещё не совсем сравнялись по форме с передними. Видимо, киты этого уровня были своеобразными «млекопитающими крокодилами» с той существенной разницей, что у ящеров позвоночник изгибается сбоку-набок, а у китов спина горбатится вверх-вниз. Протоцетусы освоили сначала морское побережье, а дальше стали заныривать всё глубже и заплывать дальше.

Ambulocetus natans
Стоило китам выйти в море – их уже было не остановить. В среднем эоцене Protocetus atavus и Aegyptocetus tarfa