Раздел I Аутэкология, или экология организмов
В 1896 г. для обозначения экологии организмов швейцарским ботаником К. Шрётером вводится термин аутэкология (от греч. autos – сам и экология). На III Международном ботаническом конгрессе (1910) аутэкология была выделена в самостоятельный раздел экологии, изучающий взаимоотношения организмов со средой, их реакции на воздействия среды. Экологические исследования быстро развивающейся науки были направлены на изучение сред, выявление влияния факторов среды на морфологические особенности, распространение, численность особей животных и растений. Итогом исследований явился огромный фактический материал о жизни особей в их взаимосвязях со средой, который послужил основой для развития других разделов экологии.
Глава 1 Организм и среда
Среди многочисленных объектов биологии особи (организму, индивидууму) принадлежит центральное место. Семьи, популяции, биоценозы взаимодействуют со средой и друг с другом только через организмы (особи). В определении В.В. Хлебовича (2002) особь есть фенотипическое проявление уникального генома, самостоятельно (автономно) устанавливающее отношения с внешней средой. Это морфофизиологическая целостная единица жизни, на которую воздействуют экологические факторы.
В первой половине XIX в. в естествознании еще господствовала концепция «организм вне среды», хотя многие ботаники и зоологи уже понимали, что растения и животные связаны со средой и находятся под постоянным ее влиянием. В 1850 г. К. Рулье, «русский предшественник Дарвина», сформулировал «закон общения животного с миром», а в 1861 г. физиолог И.М. Сеченов (1829–1905) указал, что «организм без внешней среды, поддерживающей его существование, невозможен». Биологический закон единства организма со средой называется законом Рулье – Сеченова.
В.С. Ипатов и Л.А. Кирикова (1997) определяют среду как «совокупность всех тел и веществ, потоков энергии, полей (электрического, магнитного, гравитационного), окружающих живые организмы и взаимодействующих с последними прямо или косвенно, постоянно или временно». Организм и среда представляют собой диалектическое единство, определяемое обменом веществ, в основе которого лежат противоречивые, но взаимосвязанные процессы ассимиляции и диссимиляции. Понятия «организм» и «среда» неравноправны. Как метко заметил В.П. Щербаков, среда хаотична – организм упорядочен и высокоорганизован; организм созидает – среда деструктивна; организм живет – среда мертва. В структурном, термодинамическом и информационном отношении организм неизмеримо выше среды. Организм есть сущее, а среда – условия, в которых организм существует.
1.1. Среды жизни
Организмы освоили четыре среды жизни, существенно различающиеся по специфике физико-химических условий: водную, наземно-воздушную, почву, живые организмы.
Водная среда является первой, самой древней и наиболее обширной средой жизни, занимает до 71 % площади земного шара. Жизнь в ней стала возможной задолго до образования озонового экрана и распространилась по всей ее толще до максимальных глубин (11 000 м). Водная среда более однородна, чем суша, и по разнообразию форм значительно уступает наземно-воздушной среде. В водной среде обитает около 150 000 видов животных и 10 000 видов растений.
Моря и океаны в отличие от суши не имеют физических барьеров. Они связаны между собой, что определяет широкие возможности свободного передвижения животных. Вода обладает рядом уникальных термодинамических свойств, связанных с малыми размерами молекулы, ее полярностью и наличием водородных связей (рис. 1). Она имеет особую молекулярную структуру, которая, изменяясь, способна противостоять любым внешним воздействиям – тепловым, механическим, электрическим, чем определяется своеобразие водной среды. Для воды характерна самая высокая среди жидкостей удельная теплоемкость (на нагревание 1 мл воды на 1 °C расходуется 1 кал тепла) – в 4 раза большая, чем удельная теплоемкость воздуха, что позволяет водоемам аккумулировать тепло, становиться своеобразными термостатами. Воды Мирового океана поглощают солнечной энергии почти в 2 раза больше, чем суша. Количество тепла, аккумулированного в водоемах, примерно в 21 раз превосходит годовую норму тепла, поступающего от Солнца к Земле.

Рис. 1. Молекулярная структура воды (по П. Кемпу и К. Армсу, 1980): а — молекула воды: один атом кислорода ковалентно связан с двумя молекулами водорода; б — водородные связи между молекулами воды: светлые кружки – атомы кислорода, темные кружки – атомы водорода
Благодаря высокой скрытой теплоте парообразования (при испарении 1 г воды поглощается 536 кал) водоемы не перегреваются. Они медленно нагреваются, а отдавая тепло, также медленно остывают, чем в значительной степени ослабляются годовые, суточные и даже часовые колебания их температурных режимов. Эти колебания менее резкие, чем соответствующие изменения температуры на суше. При замерзании вода уменьшает свою плотность (лед легче воды, его плотность меньше единицы), что предохраняет водоемы от сплошного промерзания, так как их поверхность покрывается льдом. Процесс замерзания сопровождается выделением значительного количества тепла, которое замедляет образование ледяного покрова.
Вода практически несжимаема. Ее состояние на дне океана мало чем отличается от состояния в поверхностных слоях. Наибольшей плотностью вода обладает при температуре 4 °C, чем обеспечивается процесс перемешивания и замерзания водных масс. Из всех жидкостей она имеет самое большое поверхностное натяжение. Благодаря силам когезии вода легко передвигается по сосудам, а мелкие организмы удерживаются или легко скользят по ее поверхности. Чем больше поверхностное натяжение, тем больше общее содержание жидкости в капиллярной системе. Вода обладает слабой испаряемостью и универсальной растеорительной способностью, в биосфере никогда не бывает химически чистой.
Специфику водной среды составляют подвижность воды, большая плотность, свет, малое содержание кислорода, температурный режим, соленость, наличие взвешенных частиц.
Подвижностью достигается перемешивание вод, выравнивание температурного режима, обеспечение гидробионтов кислородом и питательными веществами, а также в некоторой степени возможность их перемещения. По плотности вода примерно в 800 раз превосходит воздушную среду, а давление на каждые 10 м глубины возрастает на 1 атм, достигая в придонных слоях глубоководных водоемов до 1000 атм. Давление в 400–500 атм переносят головоногие моллюски, морские звезды, ракообразные и многие другие гидробионты. Плотность воды позволяет растениям со слабым развитием механической ткани и бесскелетным формам животных находиться во взвешенном состоянии, парить в воде, опираясь на ее толщу.
Лимитирующим фактором, ограничивающим распространение растений в водной толще, является свет. По мере поглощения света водой (красные лучи поглощаются у самой поверхности, а наиболее глубоко проникают сине-зеленые лучи) уменьшается световое довольствие и на глубине примерно 200 м в чистых водоемах заканчивается освещенная, или эуфотическая, зона – зона фотосинтеза (рис. 2). Глубины до 1000–1500 м занимает сумеречная, или дисфотическая, зона, а еще глубже темная, или афотическая, зона (полный мрак). При недостатке света зеленые водоросли с глубиной сменяются бурыми, содержащими кроме хлорофилла бурые пигменты фикофеин и фукоксантин, а бурые водоросли – красными, имеющими наряду с хлорофиллами а и в, каротинами, ксантофиллами специфические пигменты – красный фикоэритрин и синий фикоцианин. В зависимости от степени освещенности и спектрального состава света у водорослей меняется состав, количество пигментов и соответственно окраска. Даже при засушивании красных водорослей при недостатке света они приобретают более интенсивную окраску. Такое приспособление носит название хроматической адаптации.

Рис. 2. Примерная схема вертикальной и горизонтальной зональности моря (по X. Хилю, 1988)
Для животных, низших растений, бактерий, живущих в темной толще воды, характерно явление свечения – биолюминесценции. Генерация света происходит в результате окисления сложных органических соединений (люциферинов) с помощью белковых катализаторов (люцифераз).
Биолюминесценция характерна видам почти всех классов водных животных. В жизни животных свечение имеет сигнальное значение (ориентация в стае, привлечение особей другого пола, отвлечение), служит защитой от хищника (ослепляет его).
Освещенность водоемов зависит от количества взвешенных минеральных и органических частиц, а также от сезона года. Наиболее прозрачными считаются воды Саргассова моря (диск Секки виден до глубины 66,5 м, свет проникает на глубину до 1000 м), а прозрачность вод рек в среднем 1–1,5 м. Однако толщина слоя воды, в котором возможен фотосинтез, оказывается значительно меньше. Оптимальными для фотосинтеза являются глубины, куда доходит примерно треть солнечного света. От этой глубины интенсивность образования органических веществ в соответствии с убыванием освещенности постепенно снижается и становится минимальной на глубине, куда проникает около 1 % света. В тропических морях такая освещенность наблюдается на глубинах 40–50 м, в Балтийском море – 1—17 м. В лагунах и заливах минимальная освещенность фиксируется всего в нескольких дециметрах от поверхности воды.
Жизнь гидробионтов в воде осложняется небольшим содержанием кислорода (до 10 мл в 1 л). В атмосфере его в 21 раз больше. Основными источниками кислорода в воде являются фотосинтез и диффузия из воздуха. Диффундирует кислород очень медленно. Его молекула достигает в чистой воде глубины Юм через 11 лет, концентрация в соответствии с понижением температуры от экватора к полюсам повышается, по средним широтным данным, от 4,5–5,0 мл/л в низких широтах до 6,0–7,0 мл/л в Антарктике и до 7,5–8,0 мл/л в Арктике. Предельный минимум – 4 мл/л. Концентрация кислорода в водоемах уменьшается с глубиной, с повышением температуры, солености воды и при замерзании, что ведет к летним и зимним заморам гидробионтов. Недостатка в углекислом газе фотосинтезирующие растения не испытывают. Его в воде почти в 60 раз больше, чем в атмосфере. Содержание углекислоты, постепенно увеличиваясь с глубиной, достигает максимальных значений в придонном слое полярных областей.
Водная среда в отличие от среды наземно-воздушной характеризуется сравнительно устойчивым температурным режимом. Среднегодовая температура поверхностных слоев экваториальных вод составляет 26–27 °C, полярных вод – около 0 °C и ниже. С глубиной температура воды в океанах постепенно падает и на глубине 1000 м не превышает 4–5 °C, а на больших глубинах – относительно постоянная, колеблется от —1,8 до +2 °C. Зона между верхними слоями воды с выраженными в них сезонными колебаниями температуры и нижними слоями воды с постоянным тепловым режимом называется термоклином. Термоклин наиболее выражен в теплых морях.
Особую роль в жизни гидробионтов играет соленость воды, определяемая содержанием карбонатов, сульфатов, хлоридов и др. Количество растворенных солей в 1 л воды пресноводных водоемов не превышает 0,5 г, в морях и океанах содержится до 35 г. Так как пресноводным организмам с высоким содержанием солей (они гипертоничны по отношению к среде) и наличием проницаемых мембран постоянно угрожает излишнее обводнение в результате насасывания (рис. 3), они не потребляют воду и имеют хорошо развитую выделительную систему. Гидробионты соленых водоемов гипотоничны по отношению к водной среде. Ввиду высокого содержания солей в водной среде они, во избежание обезвоживания путем оттока воды, вынуждены постоянно пить соленую воду. По причине этих биологических особенностей пресноводные виды не могут жить в морях, а морские – не переносят опреснения. По комплексу показателей вода является уникальной средой, пригодной для жизни любых форм организмов.

Рис. 3. Осморегуляция у пресноводных (а) и морских (б) костистых рыб (из Ю. Одума, 1975)
Наземно-воздушная среда включает часть атмосферы и поверхность земли, которая служит местом прикрепления, размножения растений и животных. Воздух в отличие от воды не может обеспечить организму твердой опоры, в связи с чем наземные организмы обладают собственной опорной системой (механические ткани у растений, скелеты у животных). Опорой и одновременно источником воды и минеральных веществ для наземных растений служит почва. Малая плотность воздуха определяет его малую подъемную силу. Воздух оказывает слабое сопротивление передвижению организмов по земле, благоприятствует полету в воздухе, но затрудняет подъем по вертикали. С высотой давление воздуха понижается, обеспеченность кислородом уменьшается, что ведет к учащению дыхания у животных и их обезвоживанию. Для большинства животных высота около 6000 м составляет верхнюю границу жизни.
Среда неоднородна по горизонтали и вертикали и в отличие от океана прерывна. Постоянными физическими барьерами служат горы, реки, пустыни. Специфические особенности состоят в хорошей обеспеченности светом, постоянстве и высоком содержании кислорода (до 21 %), азота (78,1 %) и других газов, постоянном движении воздушных масс (ветер, бури, ураганы), характере осадков (ливневые или моросящие дожди, снег), а также в значительных колебаниях температурного режима. Количество тепла, как и количество выпадающих осадков, зависит от широты местности, сезона года и даже времени суток. Влажность в наземно-воздушной среде в отличие от водной сама по себе выступает важным лимитирующим фактором. С постоянными потерями воды, которые происходят при транспирации и испарении, связаны процессы миграции вещества и рассеяния энергии.
Характер распределения солнечной энергии по широте и высоте, а также неравномерность увлажнения (на суше) определяют явление географической зональности и вертикальной поясности. Годовая температура понижается при движении с юга на север на каждые 100 км (или 1° широты) на 0,5 °C, а с поднятием в горы – на 0,6 °C на каждые 100 м. Зональный характер носят основные природные процессы: климатические, гидрологические, геохимические, эдафические, геоморфологические, биогеографические и др. Закономерности и количественные характеристики природных процессов по профилю через основные природные зоны Восточной Европы отражает схема (рис. 4).
Географическая зональность представляет собой одну из важнейших природных закономерностей. Явление зональности было известно А. Гумбольдту, некоторым русским географам, но только В.В. Докучаеву предстояло в 1899 г. сказать обобщающие слова: «Вода, земля, огонь (тепло и свет), воздух, а равно растительный и животный мир, благодаря астрономическому положению, форме и вращению нашей планеты вокруг оси, несут на своем общем характере резкие и неизгладимые черты закона мировой зональности».
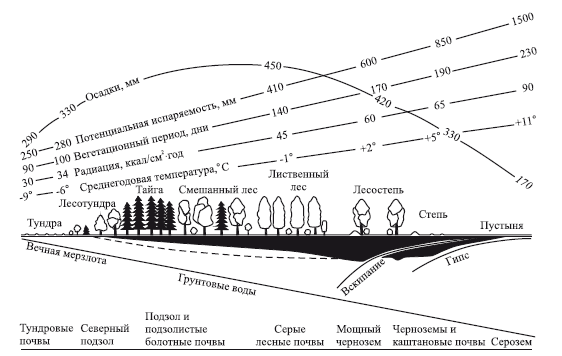
Рис. 4. Схема изменений климата, растительности и почв вдоль профиля через основные природные зоны Восточной Европы с северо-запада на юго-восток до Прикаспийской низменности. Черным показан гумусовый горизонт, штриховой линией – иллювиальный горизонт (из В. Лархера, 1978)
В соответствии с явлением зональности и поясности хорошо выражена дифференциация географической оболочки Земли, проявляющаяся в последовательной и определенной смене географических поясов и входящих в них зон. В северном полушарии выделяют естественно-исторические, или природные, зоны: ледовую, тундровую, лесотундровую, хвойных лесов, или таежную, широколиственных лесов, лесостепную, степную, полупустынную, пустынную умеренную, субтропических лесов, пустынную тропическую, саванную и экваториальных лесов. От подножья гор к вершинам, подобно широтной зональности, проявляется вертикальная поясность. Зоны и пояса отличаются определенным сочетанием тепла и влаги, характерным типом почв. Влагооборот определяет миграцию химических элементов, геохимические особенности ландшафтов, например засоленность почв пустынь, промывной режим подзолистых почв в зоне хвойных лесов. Природными особенностями зон и поясов в значительной степени определяются биоразнообразие, образ жизни и характер адаптаций организмов.
Почва как среда жизни. Образование почвы связано с литогенным процессом, ведущим к образованию глинистых минералов, и процессом биогенным, способствующим образованию гумуса – устойчивого органического вещества. Сначала влажные участки суши заселялись цианобактериальными, затем альгобактериальными и альгогрибными сообществами. Почва как корнеобитаемая система начала формироваться с появлением сосудистых растений. Она примерно 450 млн лет назад сменила древнюю кору выветривания. Для многих видов почва играла роль переходной к наземному образу жизни среды. В настоящее время она является основной средой обитания наиболее древних форм жизни на суше – прокариотов.
Почва соприкасается с подстилающей горной породой, атмосферой, поверхностными и подземными водами. Между почвой и этими природными телами, а также растениями и животными происходит постоянный обмен веществом и энергией. Почва является трехфазной системой. Твердая фаза почвы подразделяется на органическую, минеральную и органоминеральную; воздушная – на газообразную и парообразную; водная фаза почвы включает в себя воду свободную и сорбционно связанную. Твердые частицы почвы окружены воздухом и водой, поэтому она обладает рядом специфических физических свойств и существенно отличается от других сред жизни. От горных пород, на основе которых она образовалась, почва отличается особым качественным свойством – плодородием. Содержание гумуса в почве составляет масштабное отступление от правила Виноградского, согласно которому все синтезируемые биотой вещества подвергаются деструкции.
Почва обладает определенными водно-воздушным и температурным режимами и рядом специфических свойств.
Вода в почве испытывает действие сил разной природы: силы тяжести, сил молекулярного притяжения, исходящих от почвенных частиц, и сил молекулярного притяжения, действующих между самими молекулами воды. Почвенная вода находится в разных состояниях (рис. 5, 6). Вода гигроскопическая сорбируется почвой из паров, находящихся в воздухе. Она окружает почвенные частицы в виде пленки, прочно удерживается силами притяжения этих частиц, не передвигается и недоступна для использования, составляя так называемый «мертвый» запас. Кроме прочносвязанной (гигроскопической) воды в почве имеется вода рыхлосвязанная, или пленочная. Она располагается на поверхности пленки гигроскопической воды, заметно утолщая ее, может переходить от почвенных частиц с более толстой пленкой к частицам, у которых пленка тоньше (так называемое пленочное передвижение). Мельчайшие поры (диаметром 0,2–0,8 мк) занимает вода капиллярная (подвижная). Она легко испаряется, передвигается по порам (восходящий ток) и используется растениями. Наиболее крупные поры почвы заполняет вода гравитационная. Она также свободна, подвижна и передвигается вглубь почвы под влиянием силы тяжести (нисходящий ток). Дойдя до водонепроницаемого слоя, вода начинает накапливаться над ним и становится грунтовой. В почвенном воздухе содержится также вода парообразная.

Рис. 5. Типы почвенной воды
Почвенная влага представляет собой раствор. Его состав и концентрация зависят от характера почвы. Важнейшим показателем почвенной среды является реакция почвенного раствора, которая определяется соотношением концентраций свободных ионов водорода и гидроксильных ионов. Она определяет направление и скорость химических процессов. Может быть кислой (pH ниже 7), нейтральной (pH равно 7) и щелочной (pH выше 7).
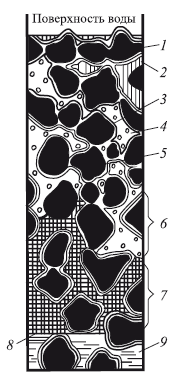
Рис. 6. Вода в почве (по Н.А. Качинскому, 1975):
1 — частицы почвы; 2 — гравитационная вода; 3, 5 – гигроскопическая вода; 4—почвенный воздух с парами воды; 6 — зона открытой капиллярной воды; 7 – зона замкнутой капиллярной воды; 8 — уровень грунтовой воды; 9 — грунтовая вода
Известковые, засоленные почвы имеют pH почвенного раствора выше 7, а сфагнумовые торфяники – ниже 4. Кислотность существенно влияет на рост растений и жизнь животных. Простейшие, например, живут в пределах pH от 3,9 до 9,7; дождевые черви не переносят pH ниже 4,4; моллюски предпочитают почвы щелочные (pH выше 7).
Для водного, воздушного и температурного режимов почвы, а следовательно, для жизни организмов, большое значение имеют сложение, структура и порозность почвы.
Сложение почвы отражает степень и характер ее плотности и порозности. Сложение определяется гранулометрическим составом почвенных компонентов, химическим составом и влажностью. Наиболее благоприятны для жизни организмов почвы с рыхлым сложением в отличие от почв плотного и очень плотного сложения. Тяжелые глинистые почвы, например, лучше удерживают влагу, но хуже прогреваются, менее воздухопроницаемы и пригодны для жизни, чем почвы песчаные.
Структура почвы определяется степенью выраженности отдельностей (агрегатов), которые имеют разную форму (кубовидную, призмовидную, плитовидную) и величину (от 1 мм до 10 см). Агрегирование определяется физическими процессами, корневой системой растений, жизнедеятельностью почвенных организмов, наличием в почве гумуса, глинистого вещества, ионов кальция и магния, гидроксидов железа и алюминия, зависит также от механического состава почвы и степени ее обработки. Структурные почвы обладают благоприятным водновоздушным режимом и максимально наполнены жизнью.
Порозность почвы, или скважность, характеризуется наличием полостей (пор) между частицами почвы и зависит от структуры почвы, ее плотности. Количество пор и их размеры уменьшаются с глубиной. Порозностью обеспечивается циркуляция воды и воздуха, жизнь и передвижение животных в почве. Благодаря наличию пор разного диаметра в почве создается обилие микросред обитания. Почва с низкой порозностью менее всего пригодна для жизни. В таких почвах кислород для большинства животных становится ограничивающим фактором. Почвенный воздух содержит всегда меньше кислорода и больше углекислоты, чем воздух атмосферный. Кислород интенсивно потребляется, а углекислый газ, наоборот, образуется и накапливается.
Количество кислорода уменьшается с глубиной, повышением влажности и температуры почвы. При пониженных температурах кислород более интенсивно заполняет пространства между почвенными частицами. Количество углекислого газа с глубиной, как правило, возрастает. При избыточном увлажнении, гниении растительных остатков в почве могут создаваться анаэробные условия, накапливаться аммиак, сероводород, метан и другие токсические газы.
Для живых организмов большое значение имеет температурный режим почвы. Основным источником тепла является солнечная радиация (прямая и рассеянная). Некоторое количество тепла почва получает от воздуха в результате разложения органических остатков и из недр земли. Резкие температурные колебания затрагивают только самые верхние ее слои и на глубине более 1 м заметно сглаживаются. Почва обладает способностью поглощать тепло, но ее теплоемкость значительно ниже теплоемкости воды.
Выражением стратиграфии почвы являются генетические почвенные горизонты. Система взаимосвязанных генетических горизонтов, производимых в ходе почвообразовательного процесса, составляет почвенный профиль. Профиль дерново-подзолистой почвы включает лесную подстилку (A0), горизонты перегнойно-аккумулятивный, или гумусовый (A1), элювиальный, подзолистый, или вымывания (А2), иллювиальный (В) и материнскую породу (горизонт Q. В горизонтах А0 и А1 происходят главным образом процессы накопления и преобразования органического вещества; в горизонте А2 преобладает вымывание; в горизонте В накапливаются минеральные вещества. Материал материнских пород постепенно преобразуется в почву.
По почвенному профилю изменяются содержание в почве гумуса и минеральных веществ, ее физические свойства, световой режим и соответственно – насыщенность жизнью. Наибольшей физической, химической и биологической активностью характеризуется верхний, гумусовый, горизонт почвы (рис. 7). В нем находится основная масса органического вещества, обитает множество живых организмов – от микроскопических бактерий и грибов, основных деструкторов органического вещества, до мелких млекопитающих (кроты, землеройки, слепыши и др.). Особое место занимают дождевые черви, которые, пропуская через пищеварительный тракт почву с органическим веществом, существенно повышают почвенное плодородие. Дождевые черви могут за год переработать на 1 га до 500 т почвы, увеличивая содержание азота в 5 раз, фосфора – в 7, калия – в 11, магния – в 3 раза.
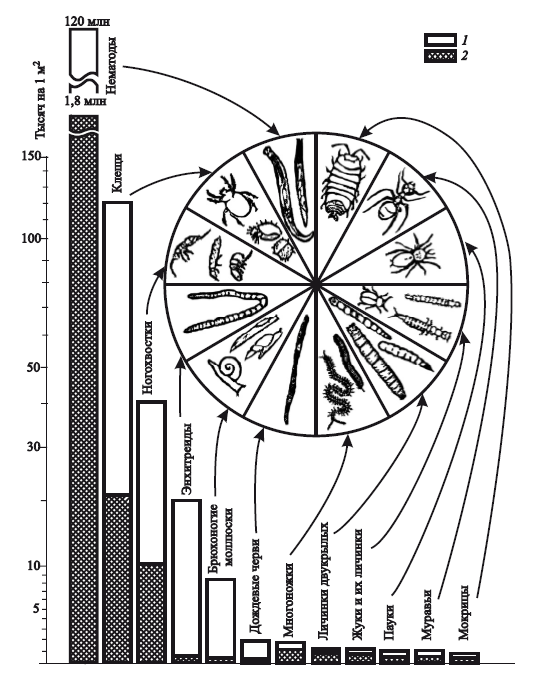
Рис. 7. Плотность населения луговой почвы (на 1 м2) до глубины около 30 см (по А. Брауну, 1988):1 — максимум; 2 — минимум
Почва – уникальная среда жизни. По данным генетиков, с ней связано 92 % генетического разнообразия. Организмы живут в порах, заполненных водой, на стенках почвенных частиц, в скважинах и целых системах проложенных ходов. Они аэробы и анаэробы, фотосинтезирующие, потребляющие и разрушающие органическое вещество. Почва, по выражению В.И. Вернадского, «сгущение жизни».
Живые организмы как среда жизни. В процессе развития живых организмов сформировалась особая биотическая среда. Предпосылками для ее возникновения послужила гомео-стазированность организмов. Явление проживания в среде организма получило широкое распространение. Даже бактерии, актиномицеты, цианобактерии служат средой для других прокариотических организмов. Среда организма отличается от других сред постоянством физико-химических параметров, в связи с чем паразит не испытывает необходимости в выработке адаптационных механизмов, что ведет к уменьшению энергетических затрат. Эта среда располагает неограниченным запасом пищи, легко доступной для усвоения, «укрывает» от потенциальных врагов, но имеет ограниченное жизненное пространство, особенно для внутриклеточных организмов, связана с трудностями обеспечения кислородом и защитными реакциями организма хозяина (более подробно о среде см. гл. 12).
Существенно отличается от сред жизни животных и растений окружающая человека среда. Она состоит из четырех неразрывно взаимосвязанных компонентов-подсистем: собственно природной среды; квазиприродной среды, возникшей в результате преобразования людьми среды природной; артеприродной среды, включающей весь вещественно-энергетический мир, созданный человеком и не имеющий аналогов в природе; среды социальной (культурно-психологический климат, создаваемый для личности, социальных групп и человечества в целом самими людьми), играющей особую роль в жизни человека.
В пределах сред жизни выделяют среды обитания и местообитания, непосредственное место жизни группы особей или вида, его «адрес». В водной среде жизни, например, различают морскую, речную, озерную среды обитания, а также среды с соленой и пресной водой, стоячей и проточной водой и др. Каждая из них располагает множеством местообитаний. По отношению к наземным животным выделяют местопребывания, называемые стацией (от лат. statio – местопребывание). Под стацией понимается часть местообитания, используемого популяцией временно либо для ограниченных целей (ночевок, питания, размножения). Популяция обыкновенной гадюки (Vipera berus[1]), например, располагает местом зимовки, местом летнего кормления и местом размножения. Стацию иногда рассматривают как местообитание популяции. Главным акцентом в понятии «местообитание» является «качество» участка: высота над уровнем моря, экспозиция, режим грунтовых вод, возможность затопления местности, физические и химические свойства почвы и др. В природе выражена иерархия местообитаний, как бы вставленных друг в друга: фитоценоз, микрогруппировка, синузия, дерево, дупло, заполненное органическим веществом. Все это разнообразие обеспечивает бесконечное множество экологических ниш и является одним из факторов многообразия видов на Земле.
Наряду с абиотической средой жизнь любой особи, популяции, вида определяется средой биотической. Жизнь организмов в косной среде привела к возникновению современной атмосферы Земли, обеспечивающей возможность кислородного дыхания живых организмов и протекания аэробных процессов, к изменению химического состава вод Мирового океана, формированию почвы, определяющей в значительной степени многообразие форм растительности на Земле. Организмы сами создали среду обитания в виде органических илов, коралловых рифов (береговых, барьерных и кольцевидных), или атоллов. Они протянулись на сотни километров в прибрежной зоне тропических морей (Большой Барьерный риф у северного побережья Австралии составляет в длину 2300 км, в ширину – от 30 до 150 км). Основными создателями рифов являются мадрепоровые кораллы (коралловые полипы) и водоросли, получающие известь непосредственно из воды, а населяют их многочисленные виды самых разных систематических групп (водоросли, губки, мшанки, кишечнополостные, оболочники, моллюски, черви, ракообразные, иглокожие, рыбы, морские змеи и др.), образуя богатейшие в мире морские биоценозы.
Значение имеет также популяционная среда, создаваемая отдельной популяцией вида. Следовательно, жизнь организмов определяется своеобразием абиотических и биотических сред, которое выражается температурным режимом и амплитудой температур, недостатком или избытком света, влаги, степенью обеспеченности кислородом, глубиной или отсутствием снежного покрова, влиянием окружающих организмов и пр. Кроме того, среды претерпевают изменения циклические (суточные, сезонные, разногодичные, влияние приливов и отливов), направленные (накопление илов, заболачивание, загрязнение, засоление), хаотические и непредсказуемые (пожары, ураганы, затопления) и др.
Весь комплекс разнообразия и сложности сред жизни, сред обитания и местообитания служит фоном естественного отбора, определяет необычайную широту и мощь этого направляющего фактора эволюции. Естественный отбор жесток, антигуманен. Он отсеивает все «неприспособленное к жизни» (отрицательный отбор), увеличивая концентрации генотипов, обладающих повышенной жизнеспособностью и плодовитостью (положительный отбор). Отбор ставит интересы вида, популяции выше «интересов особи». С одной стороны, он служит биологическому прогрессу, с другой – отбор «охраняет норму, уменьшает изменчивость», а поэтому является не только прогрессивным, но и консервативным фактором. В свою очередь, разнообразие сред, степень выраженности составляющих их компонентов обеспечивают «тонкость, дифференцированность» и «нежность» естественного отбора, определяя возможности жизни всех видов на Земле.
1.2. Факторы среды и закономерности их действия на организмы
Факторы среды – это элементы среды, оказывающие специфическое действие на организм. Влияние факторов весьма многообразно. Они выступают как раздражители, ограничители, модификаторы или имеют сигнальное значение. По своей природе факторы среды являются абиотическими, биотическими и антропическими. Особую группу составляют факторы антропогенные, действующие на человека (рис. 8).

Рис. 8. Классификация факторов среды
Действуют факторы порознь или совместно, прямо (бурелом, буревал, разнос семян) или косвенно (опосредованно). Ветер, например, увеличивает транспирацию, изменяет концентрацию углекислого газа, кислорода, иссушает среду, чем косвенно изменяет воздушный и гидрологический режимы сообщества. От высоты местности зависят температура, инсоляция, атмосферное давление. Действие факторов может проявляться в течение всей жизни или затрагивать один из этапов онтогенеза. Им обычно характерна правильная периодичность (дневная, лунная, сезонная или годовая) как прямое следствие вращения Земли вокруг оси и вокруг Солнца или смены лунных фаз. Периодичность характерна температурному режиму, освещенности, продолжительности светового дня, влажности и др., выражена и во многих биотических факторах, особенно годичных циклах. Факторы определяют расселение, географию и топографию видов, метаболизм особей, их плодовитость, продолжительность жизни, смертность, миграции, плотность популяций, вызывают у организмов сложный спектр адаптивных реакций, такие изменения, как диапауза, спячка, фотопериодизм и др.
В последнее время заметно возросла интенсивность действия антропических факторов на природные экосистемы через мелиорацию, обводнение, внесение повышенных доз минеральных удобрений, пестицидов, радиацию, физическое, химическое и биологическое загрязнения и др.
Процесс урбанизации становится мощным экологическим фактором, приведшим к возникновению таких сложных экологических проблем, как кислотные дожди, парниковый эффект, засоление и эрозия почв, утеря генофонда и биоразнообразия, деградация многих природных систем.

Ю. Либих
Одновременно урбанизация становится антропогенным фактором. На человека действуют шумы, стрессы присутствия, напряженные темпы городской жизни, а также многие факторы из группы антропических.
Закономерности действия факторов среды на организмы. Взаимоотношения организмов со средой обитания подчиняются ряду закономерностей. Первым изучать влияние факторов на рост растений начал Ю. Либих (1803–1873), немецкий химик, заложивший основы агрохимии и теории минерального питания растений. В 1840 г., задолго до возникновения экологии как науки, он заметил, что «элемент, полностью отсутствующий или не находящийся в нужном количестве, препятствует прочим питательным соединениям произвести их эффект или, по крайней мере, уменьшает их питательное действие… Отсутствие или недостаток одного из необходимых элементов при наличии в почве всех прочих делает последнюю бесплодной для всех растений».
Либих установил, что «веществом, находящимся в минимуме, управляется урожай». Такими веществами для сельскохозяйственных растений обычно являются микроэлементы. При недостатке одного из них, несмотря на хорошую обеспеченность растений светом, водой, азотом, фосфором и другими элементами, высокого урожая не бывает. Эта зависимость утвердилась в виде закона минимума Либиха.
Фактор, который в совокупном давлении среды сильнее всего ограничивает успешность жизни организма, именуется ограничивающим. Закон ограничивающих факторов был сформулирован Ф. Блэкманом в 1909 г. Знание ограничивающих факторов имеет большое практическое значение. Агротехническим приемом можно изменить состояние среды, снять или ослабить действие ограничивающего фактора, что дает возможность повысить урожай, качество продукции.
Ограничивающим может быть любой из факторов. В водной среде, в зависимости от ее глубины, им является кислород. Жизнь наземных растений и животных чаще всего лимитируется температурным фактором. Для урожая сельскохозяйственных, плодово-ягодных растений критическим становится погодный режим, при котором массовое развитие получают вредители и возбудители болезней, например фитофтора (Phytophthora infestans); урожай абрикоса обыкновенного (Armeniaca vulgaris) в Беларуси лимитируется весенними заморозками всего в 1–2 °C во время цветения, хотя растения вида способны переносить в состоянии покоя морозы до 25–28 °C.
Уточняющим дополнением к этому закону послужил закон компенсации (взаимозаменяемости) факторов Рюбеля: один экологический фактор может быть компенсирован другим близким фактором. Например, недостаток кальция при построении раковины некоторые моллюски заменяют использованием стронция. Видимо, подобная компенсация весьма относительна. Этому закону противостоит закон незаменимости фундаментальных факторов Вильямса, который считал, что свет, воду, а также углекислый газ, азот, фосфор, многие микроэлементы нельзя заменить другими факторами. Общеизвестно, что при отсутствии энергии нет жизни, как и при полном безводье.
В. Шелфорд, изучая жуков-скакунов, установил, что жизнь этих насекомых протекает в очень точных пределах факторов среды. Кладка яиц производится под камни, где мало света, в песчаную, рыхлую, хорошо дренируемую, с небольшим содержанием гумуса почву при очень узком интервале температур. Только в таких условиях выживают личинки. Эти и другие «точности» в жизни животных привели Шелфорда к понятию «пределы выносливости», а затем к формулировке закона толерантности. Закон гласит, что лимитирующим фактором процветания организма (вида) может быть как минимум, так и максимум экологического воздействия, диапазон между которыми определяет величину выносливости (толерантность) организма к данному фактору. Схематически содержание закона отражает рис. 9. Возрастание напряженности фактора, начиная с предельно малого значения, вызывает сначала повышение эффекта, а затем резкое понижение и гибель. В соответствии с этим в пределах диапазона выносливости имеются зоны: оптимума, или комфорта, где вид процветает; пессимума, или минимума, в которой вид более редок, жизненность особей пониженная. За пределами выносливости, фиксируемыми минимальной и максимальной кардинальными (критическими) точками, особи вида погибают.

Рис. 9. Пределы выносливости вида в зависимости от интенсивности фактора
Закон толерантности стал основой многочисленных исследований, которые позволили установить пределы существования для многих видов растений и животных, выявить закономерности их распределения в природе. Степень способности вида выдерживать действие фактора среды получила название экологической валентности (от лат. valentia – сила). Степень толерантности видов к определенному фактору среды очень разная. Экологическая валентность может меняться в соответствии с возрастным состоянием особи. К засухе, например, наиболее чувствительны всходы. Особенно критическим в жизни организмов является период размножения. Пределы толерантности для размножения особей, прорастающих семян, яиц, эмбрионов, проростков, личинок обычно значительно уже, чем для взрослых растений и животных. Тополь белый, тополь черный, например, растут на разных по механическому составу и влажности почвах, но семена прорастают только на увлажненной, незадерненной, хорошо аэрируемой почве.

Рис. 10. Относительные пределы толерантности стенотермных (I, III) и эвритермных (II) организмов (по Рутнеру, 1953)
По отношению к температурному фактору различают виды высокой валентности – эвритермные (от греч. eurys – широкий) и низкой валентности – стенотермные (от греч. stenos – узкий, тесный) (рис. 10). Эвритермными видами являются пшеница (Triticum durum), люцерна серповидная (Medicago falcate), клевер луговой (Trifolium pretense). Пшеница растет в пределах температур 0—42 °C, люцерна и клевер – 1—37 °C. Развитие колорадского жука протекает в более узком интервале температур – 12–33 °C. При более высоких температурах развитие личинок приостанавливается. Оптимум развития составляет температурный режим 25–30 °C. Выраженным стенотермным видом является шоколадное дерево (Theobroma cacao). Деревца погибают при температуре 10 °C, угнетены при 15 °C, наиболее успешно развиваются при температуре 18–20 °C. Крайне стенотермным видом является антарктическая рыба пестряк (Trematomus bernacchii). Она живет в интервале температур от —2 до +2 °. По отношению к засолению среди животных выделяют виды эвригалинные и стеногалинные (от греч. hals – соль). Эвригалинные животные могут существовать при солености воды от 0,5 до 250%о, а виды стеногалинные, наоборот, обитают только в узком диапазоне изменений солености воды.
Наряду с видами, которые имеют широкий диапазон толерантности к одному фактору, но узкий к другому, существуют виды с широким диапазоном толерантности ко многим факторам. Эти виды имеют широкую экологическую амплитуду и называются эерихорными (от греч. chora – место, пространство), или эерибионтными. Эврихором является сосна обыкновенная (Pinus sylvestris), вид с ареалом, в пределах которого она успешно растет в разных климатических условиях (распространена от Шотландии и гор Андалузии в Испании до Охотского моря в Азии, встречается в горах Крыма и Кавказа) на почвах дерново-подзолистых, подзолистых, торфяно-болотных, разного механического состава и разной трофности, включая все типы леса, начиная от сосняка лишайникового до сосняка сфагнового. Серая крыса обитает в любых условиях, размножается при отрицательных температурах (промышленные холодильники) и при температуре до +55 °C (обшивки паровых котлов), передвигается на большие расстояния вдоль железных дорог, питается всеми видами корма.
Стенохорные (стенобионтные) виды имеют узкую экологическую амплитуду, обитают в специфических условиях (пещеры, горячие источники). Чаще являются эндемиками, паразитами. Произрастание ели сербской (Picea omorica) приурочено к известняковым отложениям крутых склонов гор Балканского полуострова. Эндем Восточного Закавказья (хребет Ейлар-Оухи) сосна эльдарская (Pinus eldarica) растет исключительно в зоне сухой степи на засоленных и известковых почвах.
На организмы действуют не отдельные факторы, а их совокупность. Действие одного фактора зависит от уровня выраженности прочих факторов. Корреляции с одним фактором не бывает. В 1909 г. немецкий агроном и физиолог А. Митчерлих показал, что величина урожая зависит от совокупности одновременно действующих факторов, что нашло выражение в законе эффективности факторов. В 1918 г. Б. Бауле закон переименовывается в закон совокупного (совместного) действия факторов, а затем получает окончательное название закон Митчерлиха – Бауле. При совместном действии происходит взаимообмен между факторами, их усиление (явление синергизма) или ослабление. Например, сильный ветер увеличивает транспирацию и испарение; высокие температуры животными переносятся труднее при повышенной влажности. Синергическое действие хорошо проявляется при правильном подборе комплекса удобрений.
Знание отмеченных законов помогает решать ряд практических задач, создавать, например, оптимальные условия при выращивании растений, содержании животных. При этом необходимо правильно выделять наиболее существенные факторы в жизни организмов, определять их функционально важную роль на основных этапах онтогенеза. При решении практических проблем применим закон фазовых реакций («польза – вред»), согласно которому малые концентрации токсиканта действуют на организм в направлении усиления его функций (их стимулирования), а более высокие, наоборот, угнетают или смертельно токсичны. Однако эта закономерность справедлива не для всех токсикантов и особенно спорна для малых доз радиации.
Глава 2 Адаптивная морфология организмов
Существование организмов в разнообразных и постоянно меняющихся условиях сред жизни возможно только благодаря сложным адаптивным программам, в основе которых лежат разные механизмы. Способность к адаптациям составляет одно из главных свойств живой материи. Адаптации возникают и развиваются одновременно с возникновением и становлением вида и проявляются на разных уровнях организации живой материи.
2.1. Адаптации организмов, правила и механизмы
Адаптация (от позднелат. adaptatio – приспособление) – это процесс приспособления строения и функций организмов (особей, видов) и их органов к факторам среды. А.С. Северцов (1981) определяет адаптацию как любое изменение организации, снижающее смертность под воздействием факторов среды. Адаптивным является всякий признак, снижающий элиминацию особей. В процессе адаптации возникает соответствие между факторами среды и способностями организмов процветать в ней, развивается гармония между организмом и средой.
Адаптации развиваются в ходе эволюции видов, в процессе адаптациогенеза. Естественный отбор объективно определяет направление возникающих адаптаций, творчески выбирая из разнообразного биологического материала (фенотипов) лишь тот, который оказывается пригодным для данных условий. Адаптации являются результатом жизни и смерти многих предшествующих поколений. Отсутствие отбора по признаку ведет к его разрушению и исчезновению. Адаптации всегда носят относительный характер и развиваются на основе определенных онтогенетических предпосылок или адаптационных явлений, которые наследственно не закреплены. Такие онтогенетические (фенотипические) адаптации обратимы, но наряду с генотипическими играют также важную роль в жизни организмов, и чем выше они стоят на эволюционной лестнице, тем их роль больше.
В сложных условиях среды каждый организм, популяция существуют благодаря комплексу адаптаций генотипических и онтогенетических, которые действуют одновременно, не исключая друг друга. Согласно правилу экологической индивидуальности Л.Г. Раменского, каждый вид адаптирован по-своему, индивидуально. В составе кактусов, например, имеются стеблевые (высокие древовидные, кронообразующие или канделябровидные, колонновидные, кустарниковидные) и карликовые формы. Карликовые кактусы являются шаровидными, ежеобразными, шишковидными и др. Адаптация вида совершается к определенной, строго специфической экологической нише, что отражает экологическая аксиома Дарвина. Высокая адаптированность к одному фактору, по закону относительной независимости адаптации, не дает такой степени приспособленности к другому фактору, она может даже ограничивать их действие. Организм, адаптированный, например, к водной среде, не может жить на суше.
В энергетическом отношении адаптация заключается в способности поддержания энергетического баланса организма, что было впервые аргументировано Н.И. Калабуховым. Энергетический баланс животные поддерживают путем снижения затрат энергии или, наоборот, повышенным ее потреблением. По первому пути пошли пойкилотермные животные, некоторые млекопитающие и птицы. Они обладают способностью избегать неблагоприятного действия внешних условий. Большинство гомойотермных животных активно противостоят экстремальным воздействиям среды, что сопряжено с колоссальными добавочными энергетическими расходами.
Адаптации представлены бесконечным разнообразием, трудно поддающимся обобщению. В основе этого разнообразия лежат следующие механизмы: генетические, физиологобиохимические, анатомо-морфологические, поведенческие, онтогенетические.
Генетические механизмы. Одной из ответных реакций организмов на неблагоприятные условия среды, чаще всего на суровые климатические условия, является полиплоидия. Полиплоидные формы, как правило, крупнее и способны существовать в условиях сурового климата. Больше всего полиплоидных видов растений отмечено в высокогорьях и в Арктике. Их количество увеличивается в широтном (с юга на север) и меридиальном (с запада на восток) направлениях по мере ухудшения климатических условий. Устойчивость полиплоидов объясняется увеличением объема генетической информации (числа геномов и генов), в результате чего возрастает уровень рекомбинационной изменчивости.
Регуляцией активности генов (выключение одних генов и активация других), отвечающих за ферменты в клетках, катализирующие одни и те же реакции, но при различных температурах, достигается акклимация, или процесс тепловой закалки организмов. Акклимация реализуется обычно за 10–15 суток, не ведет к морфологическим преобразованиям и протекает, например, при интродукции видов.
Физиолого-биохимические механизмы. Основой этих механизмов является гомеостаз внутренней среды организма. В ответ на возмущения во внутренней среде, пришедшие из среды внешней, нервной или нервно-гуморальной системой запускаются разные механизмы. Терморегуляцию гомойотермного животного отражает схема (рис. 11). У гомойотермных животных в ответ на снижение температуры окружающей среды происходит, например, рефлекторное усиление теплопродукции. Оно достигается путем выделения тепла функционирующими органами, тканями (химическая терморегуляция) и специфическим терморегуляторным теплообразованием (сократительный термогенез). Сократительный термогенез протекает в скелетной мускулатуре, не затрагивая ее прямую моторную деятельность, а также в покоящейся мышце. У млекопитающих одной из форм термогенеза является также окисление особой бурой жировой ткани, содержащей большое количество митохондрий и пронизанной многочисленными кровеносными сосудами. Под действием холода увеличивается кровоснабжение бурого жира, интенсифицируется его дыхание, возрастает выделение тепла, от которого нагреваются в первую очередь вблизи расположенные сердце, крупные сосуды, лимфатические узлы, а также центральная нервная система.

Рис. 11. Схема терморегуляции гомойотермного животного
Регуляторные реакции при перегреве организма представлены также различными механизмами усиления теплоотдачи во внешнюю среду и отмечаются при первых признаках начинающегося перегрева. Ими являются интенсификация испарения влаги с поверхности тела, верхних дыхательных путей, потовыделение. У растений защитой от перегревания служит процесс транспирации, который из-за большого расхода воды называют «неизбежным злом». При потреблении 2000 т воды усвоенная и трансформированная вода составляет всего 3 т (0,15 %) (рис. 12). Уменьшение потерь воды при транспирации достигается разными приспособлениями. Сложными физиологическими процессами (кислотный метаболизм толстянковых, С4-фотосинтетический метаболизм) обеспечивается возможность жизни ряда растений в аридных условиях, а у С4-растений, кроме того, достигается очень высокая продуктивность в самых экстремальных условиях Земли, в «зонах экстраинсоляции» (пустыни Аравийская, Каракумы, Гоби, Сахара и др.).
Анатомо-морфологические механизмы выражаются в разнообразных морфологических и анатомических приспособлениях, которые формируются в соответствии с образом жизни в определенной среде и выполняют защитную функцию в межвидовых отношениях. Спектр их весьма разнообразен:
• твердые покровы, своеобразные защитные образования в виде панцирей: хитиновый покров членистоногих; прочные раковины моллюсков; ороговевшие покровы (щитки, чешуи, шипы, панцири) у рептилий; богатые кремнеземом оболочки клеток, кутикула, восковой налет на листьях и побегах растений;
• защитные образования и выделения: войлочное, паутинистое и железистое опушения; твердые выросты эпидермиса (шипы) у растений; иглы, колючки, воскообразная слизистая пленка и др.;
• ткани: эпидермис, пробка, палисадная ткань и др.;
• форма, размеры органов, тела животных и растений; форма и размеры устьиц, листовых пластинок, корневых систем; жизненные формы, нанизм и др.;
• приспособительная окраска (покровительственная, расчленяющая, предупреждающая, отпугивающая); мимикрия (миметизм и мимезия).

Рис. 12. Расход воды на фотосинтез (по X. Пенмэну, 1972)
Поведенческие механизмы. В основе поведенческих реакций (поведения выживания) главным является способность организмов к перемещению в пространстве. Чем активнее животное, тем больше оно получает экологической информации и тем реже прибегает к пассивным способам защиты. Обычная реакция особей на воздействие среды выражается в способности зарыться, убежать, уплыть, улететь, а также в виде фото- и геотаксисов. Кроме адаптивного перемещения в пространстве (миграции, перелеты), реализующего стремление возвратиться в нейтральные или оптимальные условия, некоторым животным характерен так называемый, по определению А.Г. Гинецианского, изолирующий рефлекс. Многие животные обладают способностью затаиться, укрыться в убежище. Двустворчатые моллюски, усоногие ракообразные от вредного воздействия изолируются, например, смыканием створок раковины. К поведению выживания относится также полное обездвижение в ответ на появление хищника. «Поза покоя» характерна насекомым, рыбам, амфибиям, птицам, не обладающим средствами активной защиты от врагов. Эффект поведенческой реакции увеличивается, если он сочетается с покровительственной окраской. Большая выпь (Botaurus stellaris), например, в позе настороженности мало заметна даже на близком расстоянии.
Комплексы поведения проявляются стереотипно, быстро и сходно у разных особей одного вида с минимальными затратами энергии. Они создаются естественным отбором, как и другие генетические программы, и адаптивны в среде, которая была сравнительно постоянной на протяжении истории вида. На основе видового стереотипа в процессе онтогенеза вида и индивидуального опыта особей вырабатываются поведенческие реакции лабильного типа, ориентированные на разные сигналы реальной ситуации.
Онтогенетические механизмы направлены на ускорение или замедление индивидуального развития. У малолетников ускорение онтогенеза достигается неотеническими модификациями – появлением, например, репродуктивных органов уже на ювенильной стадии развития организма. Повышение устойчивости организмов обеспечивается понижением жизнедеятельности, падением уровня обмена веществ, ослабеванием или даже остановкой отдельных жизненных функций, что выражается в наступлении покоя. Такое состояние является обязательным этапом онтогенеза в нестабильных условиях среды обитания (жаркое сухое лето, суровая зима). Покой бывает вынужденным (гипобиоз), наступая сразу под действием фактора, что ведет, например, к оцепенению некоторых насекомых, и физиологическим (криптобиоз), связанный с длительной обязательной физиологической подготовкой при наступлении этого состояния и при выходе из него. Криптобиоз у разных организмов протекает по-разному: покой семян, спор, цист; глубокий покой растений; спячка животных в жаркое сухое и зимнее холодное время; диапауза. Диапауза – состояние покоя, сопровождающееся задержкой роста и развития на разных этапах онтогенеза: от яйца до имаго включительно. Куколки бабочки-желтушки, эндема Новой Земли, могут зимовать, например, несколько лет подряд, дожидаясь более теплого сезона. Процессы развития тормозятся железами внутренней секреции, вырабатывающими гормоны. У видов с простым и упрощенным строением возможна полная временная остановка жизни – анабиоз.
Все рассмотренные адаптационные механизмы в общем отражают приспособления организмов к факторам среды. Для более глубокого понимания их действия следует учитывать, что адаптации проявляются, как отмечалось выше, на всех уровнях организации материи. Совместное функционирование разных по уровню адаптивных систем, которые действуют одновременно и не переходят друг в друга, обеспечивает максимальную эффективность приспособления вида к конкретным условиям (рис. 13–15). По отношению к интенсивности освещения, например, приспособления растения на клеточном уровне проявляются в гранальной структуре, количестве, размерах, положении и поведении хлоропластов, количестве и формах хлорофилла, толщине клеточной оболочки, концентрации клеточного сока и др. Особую роль на тканевом уровне играет строение листа, степень выраженности его световой или теневой структуры, наличие кутикулы, воскового налета, опушения. Реакция листа как органа растения на свет выражается в его форме, размерах, способности менять положение, а дерева как организма – в выработке индивидуальной адаптивной архитектоники кроны (размеры, форма, плотность), световой мозаики. Адаптивное значение имеют и поведенческие реакции единичной особи.

Рис. 13. Адаптации растений к свету на клеточном уровне. Схема изменения положения хлоропластов и хроматофоров в клетках растений при разной степени освещенности (из T.K. Горышиной, 1979): а — ряска; б — моугеотия; в — селагенелла; 1 – слабый свет; 2 — сильный свет; 3 — темнота

Рис. 14. Адаптации растений к свету на тканевом уровне. Световая структура листа акации песчаной (по А.П. Шенникову, 1950)

Рис. 15. Адаптации растений к свету на организменном уровне. Листорасположение у подроста липы мелколистной в разных условиях освещения (из Т.К. Горышиной, 1979): а — под пологом леса; б— на открытом месте
Адаптации на популяционном уровне выражаются в гетерогенности особей популяции по отношению к свету, что способствует расширению границ местообитания. Их носителями являются группы особей, например особи молодые и генеративные. Экосистемными адаптациями определяется положение популяции в экосистеме по отношению к другим популяциям, что достигается пространственной структурой системы.
На разных уровнях разнообразны также адаптации растений к низким температурам. На клеточном уровне они заключаются в структурном изменении клеток. Клетки становятся менее вакуолизированными, цитоплазма уплотняется, увеличивается вязкость, возрастает жесткость клеточных стенок. В клетках увеличивается количество сахаров – основных источников энергии, аминокислот, нуклеиновых кислот, белков и липидов. Процессы в клетках инициируют межклеточное льдообразование. Кристаллы в межклетниках «вытягивают» воду из протопластов к центрам кристаллизации и подогревают их выделяемым теплом. Цитоплазма обезвоживается, клетки уменьшаются, а межклетники увеличиваются в объеме. На организменном уровне смягчение действия морозов достигается синхронизацией сезонного ритма роста растений и сезонного хода температур, местоположением растения, поглощением тепла из окружающей среды, метаболическим обогревом (активность дыхания на холоде заметно возрастает), теплопродукцией в процессе замерзания воды на поверхности растения, подстилки и др.
Согласно принципу биологического эпиморфизма В.Н. Новосельцева, любая адаптация тем эффективнее, чем большим числом параллельных механизмов она поддерживается. Например, растения затененных местообитаний преодолевают неблагоприятные условия посредством таких сходных адаптаций, как формирование листовой мозаики, утончение листа, изменение состава пигментов, увеличение количества хлорофилла и др.
2.2. Анатомо-морфологические адаптации
Среди множества адаптаций особенно разнообразны адаптации анатомо-морфологические, представленные разнообразием экологических групп и жизненных форм.
22.1. Экологические группы
Попытки классификации организмов по отношению «к среде в конкретной обстановке» предпринимались еще во второй половине XVIII в. Наиболее успешно разрабатывались классификации растений. Аналогичные работы с животными были менее плодотворными в связи с очень большим разнообразием и проживанием во всех средах жизни. В XIX в. Скау по отношению к свету различал растения светолюбивые, тенелюбивые и растения, обитающие в темноте. Унгер классифицировал растения по отношению к почве, Декандоль – по отношению к теплу. С введением Р. Дювинью термина «экологические группы» начала складываться система экологической классификации организмов, ставшая основой развития аутэкологии. В каждую экологическую группу объединяются виды с более или менее сходными потребностями к определенному фактору среды. Принято различать, например, экологические группы организмов по отношению к свету, влаге, теплу и другим факторам. В зависимости от этапа онтогенеза положение организма в экологической системе может меняться.
В основу экологической классификации животных в связи с их разнообразием и обитанием в разных средах положены различные критерии: отношение к факторам среды, образ жизни, способ передвижения и даже размеры особей.
Среди водных животных (гидробионтов) выделяют группы видов по месту обитания в толще воды и специфике передвижения в ней: пелагические виды, бентос и перифитон. Пелагические виды (обитатели толщи воды – пелагиали) представлены нектоном (активно передвигающиеся виды) и планктоном (виды, парящие в толще воды благодаря ее высокой плотности). Образ жизни определяет особенности их приспособлений. Виды нектона (от греч. nektos – плавающий), включающего рыб, ластоногих, головоногих моллюсков, китов, в пресноводных водоемах – земноводных, крупных насекомых, являются активными пловцами. Некоторые могут развивать скорость до 110 км/ч и занимают обширные ареалы. Они имеют обтекаемую, торпедообразную форму тела, развитую мускулатуру, хорошо развитые органы передвижения. Непарные плавники рыб, расположенные вдоль средней линии тела, помогают стабилизировать тело, а парные (грудные и брюшные) используются для руления и балансирования и вместе с хвостовым плавником обеспечивают продвижение организма вперед через толщу воды. Уменьшение трения между телом и водой достигается смазыванием чешуи выделениями кожных желез. По биомассе первое место в составе нектона занимают рыбы (80–85 %). На долю головоногих моллюсков приходится 10–15 %, млекопитающих – менее 5 % от общей биомассы нектона. Планктон (от греч. planktos – парящий), занимающий ключевую позицию в водной экосистеме, включает наряду с мелкими животными (зоопланктон) и растения (фитопланктон – одноклеточные и колониальные водоросли). Видам этой группы не характерна способность активного передвижения (кроме вертикальных миграций), и все их приспособления направлены на повышение плавучести (рис. 16). Плавучесть достигается путем увеличения относительной поверхности тела (сплющенность, выросты), уменьшения размеров и плотности тела (редукция скелета, накопление капелек жира, наличие пузырьков воздуха, воздухоносных камер и др.).

Рис. 16. Увеличение относительной поверхности тела у планктонных организмов (по С.А. Зернову, 1949). Расчлененные формы: а — личинка берегового морского ежа; 6—личинка многощетинкового червя нереис; в — личинка береговой улитки; г – медуза; д — веслоногий рачок
Особую группу в составе планктона составляет ней стон (от греч. neustós – плавающий), виды которого населяют пленку поверхностного натяжения воды. Эти виды активно передвигаются, скользя по водной поверхности, некоторые ведут неподвижный образ жизни. Составляют группу простейшие и другие микроорганизмы, насекомые. Наиболее обычны жуки-вертячки, клопы-водомерки и др.
Гидробионты, обитающие на дне и в толще осадков водоемов, составляют бентос (от греч. benthos – глубина). Группа также очень разнообразна. В нее, кроме животных (зообентос), включают бактерии и водоросли (фитобентос), которые получают развитие при небольшой толще воды и некотором освещении. Виды бентоса составляют прикрепленные или медленно передвигающиеся организмами, многие живут в толще органического вещества, являются роющими животными. Среди них есть фильтраторы (например, двустворчатые моллюски) и грунтоеды (например, брюхоногие моллюски).
Особую экологическую группу составляет перифитон. Включает виды (животные, а также растения), прикрепленные к стеблям, листьям растений и разным другим поверхностям в толще воды.
По отношению к воде животные классифицируются на гидрофилы (животные, постоянно живущие в воде), гигрофилы (влаголюбивые наземные животные, живущие на заболоченных территориях, в поймах рек, а также в почве), мезофилы (наземные, наземно-воздушные животные мезофитных местообитаний) и ксерофилы (животные засушливых местообитаний). Ксерофильные виды (насекомые, грызуны) не пьют, используют метаболическую воду, распространены преимущественно в полупустынях, пустынях и степях, имеют водонепроницаемые покровы и способны довольствоваться метаболической водой.
По отношению к температурному фактору животные разделяются на эвритермные и стенотермные виды (в соответствии с широкой и узкой амплитудой температур жизнедеятельности). Эвритермными являются преимущественно животные умеренного пояса. Среди стенотермных животных различают теплолюбивых (термофильных) и холодолюбивых (термофобных, или криофилов). Термофобные виды сравнительно немногочисленны, приурочены к регионам с суровым климатом (арктические и антарктические пустыни, тундры, высокогорья, морские глубины).
В связи со сложностью почвенной среды (большая суммарная поверхность почвенных частиц, обилие мелких полостей и скважин, заполненных водой или воздухом), занимающей промежуточное положение между водной и наземной средами, мир почвенных организмов необычайно экологически разнообразен. В его составе – виды разных систематических групп и разной организации, включая одновременно микроскопических одноклеточных простейших и млекопитающих. В основу классификации почвенной фауны положены размеры особей. Величина организмов лимитируется физическими особенностями почвенной среды. С величиной в значительной степени связаны характер передвижения, местоположение, образ жизни и особенности функционирования животных.
В полостях и скважинах, заполненных водой, живут фактически водные «плавающие» одноклеточные виды (амебы, жгутиковые, инфузории) и микроскопические мелкие беспозвоночные (коловратки, нематоды). Для некоторых из них, а также для многих микроорганизмов средой обитания является пленочная влага, окружающая почвенные частицы. Наиболее мелкие виды почвенной фауны объединяются в микрофауну. Более крупные организмы (от долей до 2–3 мм), в основном членистоногие (клещи, насекомые), корненожки, составляют мезофауну. Они подвижны, живут в полостях, заполненных воздухом, насыщенным водяными парами, получая воздух через тонкие покровы тела. С изменением влажности воздуха виды мигрируют в глубь почвы для защиты от высыхания. При пересыхании почвы некоторые виды способны образовывать цисты-капсулы с плотными оболочками и переносить в течение длительного времени неблагоприятные условия. Для видов макрофауны (многоножки, мокрицы, ногохвостки, дождевые черви) почва является плотной средой, оказывающей значительное механическое сопротивление при движении. В связи со сравнительно крупными размерами (2—20 мм и более) виды не могут пользоваться естественными полостями почвы. Они расширяют их или роют собственные ходы.
Почвенная среда определяет форму тела, морфологические особенности и образ жизни этих животных. Тело их тонкое, вытянутое, способное изгибаться в узких извилистых ходах почвы. Одни из них (например, дождевые черви) способны за счет сокращения мускулатуры кожно-мускульного мешка изменять диаметр тела. Сначала дождевой червь, зафиксировав задний конец тела, утончает и удлиняет передний, проникая в узкую почвенную щель. Овладев ею, он фиксирует переднюю часть тела, где создается сильное гидравлическое давление, за счет которого щель расширяется и обеспечивается поступательное продвижение. Вертикальные миграции дождевых червей достигают иногда 3 м. Ряд животных (личинки насекомых, многоножки), зафиксировав тело (подпорки, зубцы, крючки), проделывают ходы путем разрыхления почвы челюстями или передними конечностями, расширенными и укрепленными, с последующим ее отгребанием и закупоркой хода специальными приспособлениями на заднем конце тела, называемом «тачкой». Закрытием хода обеспечивается постоянство среды.