Что такое жизнь?
На основе лекций, прочитанных при содействии Дублинского института перспективных исследований в Тринити-колледже, Дублин, в феврале 1943 г.
Памяти моих родителей
Предисловие
В начале 1950-х годов, будучи молодым студентом-математиком, я мало читал, но уж если читал, то в основном Эрвина Шредингера. Мне всегда нравились его работы, в них чувствовалось волнение открытия, сулившее действительно новое понимание загадочного мира, в котором мы живем. В этом смысле особенно выделяется короткая классическая работа «Что такое жизнь?», которую, как я теперь понимаю, непременно следует поставить в один ряд с самыми влиятельными научными трудами XX века. Она является мощной попыткой осознать настоящие тайны жизни – попыткой, сделанной физиком, чьи собственные проницательные догадки сильно изменили наше представление о том, из чего состоит мир. Мультидисциплинарность книги была необычной для своего времени, однако она написана с подкупающей, хотя и обезоруживающей скромностью на уровне, доступном неспециалистам и молодым людям, стремящимся к научной карьере. На самом деле, многие ученые, внесшие фундаментальный вклад в биологию, такие как Б. С. Холдейн[1] и Фрэнсис Крик[2], признавали, что на них оказали значительное влияние различные идеи, пусть и спорные, выдвинутые в этой книге вдумчивым физиком.
Как и многие другие работы, повлиявшие на человеческое мышление, «Что такое жизнь?» излагает точки зрения, которые, будучи усвоенными, представляются почти самоочевидными истинами. Тем не менее их по-прежнему игнорирует множество людей, кому следовало бы понимать, что к чему. Как часто мы слышим, что квантовые эффекты не имеют особого значения в биологических исследованиях или даже что мы потребляем пищу, чтобы получить энергию? Данные примеры подчеркивают непреходящую значимость книги Шредингера «Что есть жизнь?». Без сомнения, ее следует перечитать!
Введение
Предполагается, что ученый обладает полным и всеобъемлющим знанием о вещах, полученным из первых рук, а следовательно, не должен писать о том, в чем не является экспертом. Как говорится, noblesse oblige[3]. Сейчас я попрошу вас забыть про noblesse, если таковое имеется, и освободиться от соответствующих обязательств. Я оправдываю это следующим образом: от наших праотцев мы унаследовали сильное стремление к единому, всеохватывающему знанию. Само название высших образовательных учреждений напоминает нам, что с античных времен и на протяжении многих столетий наибольшее внимание уделялось аспекту универсальности. Однако рост – в ширину и глубину – различных ветвей знания в последние сто с небольшим лет заставил нас столкнуться со странной дилеммой. Мы отчетливо ощущаем, что лишь начинаем собирать надежный материал, из которого можно вывести общую сумму всех известных вещей. Но с другой стороны, теперь отдельный ум способен одолеть только небольшой, специализированный фрагмент знания.
Я вижу лишь один способ справиться с этой дилеммой (иначе наша истинная цель будет утрачена навеки): кто-либо должен взять на себя синтез фактов и теорий, даже полученных из вторых рук и неполных, рискуя выставить себя глупцом.
Таково мое оправдание.
Не следует недооценивать языковые сложности. Родной язык – как скроенная по фигуре одежда, и человек чувствует себя неуютно, когда лишается доступа к нему и вынужден пользоваться другим языком. Я хочу выразить благодарность доктору Инкстеру (Тринити-колледж, Дублин), доктору Патрику Брауну (колледж Святого Патрика, Мейнут) и – последнему по счету, но не по значению – мистеру С. К. Робертсу. Им было нелегко подогнать под меня новую одежду и убедить отказаться от «оригинальных» оборотов. Если часть их пережила редактуру моих друзей, это моя вина.
Заголовки разделов изначально должны были представлять собой краткое содержание, и текст каждой главы следует читать in continuo[4].
Менее всего свободный человек размышляет о смерти. В своей мудрости он размышляет не о смерти, а о жизни.
Глава 1 Классический физический подход к предмету
Я мыслю, следовательно, существую.
Эта небольшая книга родилась из цикла публичных лекций, прочитанных физиком-теоретиком перед аудиторией из четырехсот человек, которая не сократилась даже после изначального предупреждения о сложности предмета и о том, что лекции нельзя назвать популярными, хотя в них практически не используется самое ужасное оружие физика, математическая дедукция, – не потому, что данный предмет можно объяснить без привлечения математики, а просто он слишком запутан для полного математического описания. Другой особенностью, которая придавала лекциям некий популярный оттенок, было намерение лектора объяснить и биологам, и физикам фундаментальную идею, лежащую на стыке биологии и физики.
В действительности, несмотря на разнообразие затрагиваемых тем, затея призвана донести лишь одну мысль – маленький комментарий к большому и важному вопросу. Чтобы не заблудиться, составим короткий план.
Большой, важный и весьма обсуждаемый вопрос заключается в следующем:
Как физика и химия объясняют события в пространстве и времени, происходящие в пространственных рамках живого организма?
Предварительный ответ, который попытается установить и обосновать эта книга, можно кратко изложить так:
Очевидная неспособность современных физики и химии объяснить подобные явления вовсе не означает, что эти науки не могут их объяснить.
Данное замечание было бы весьма тривиальным, если бы единственным его предназначением являлось пробудить надежду на достижение в будущем того, чего не удалось получить в прошлом. Однако его значение намного более оптимистично: эта неспособность имеет подробное объяснение.
Сегодня, благодаря блестящей работе биологов, в основном генетиков, за последние тридцать-сорок лет, мы знаем достаточно о действительной материальной структуре организмов и об их работе, чтобы заявить и назвать точную тому причину: современные физика и химия не могут объяснить пространственно-временные события, происходящие в живом организме.
Взаимодействия атомов в жизненно важных частях организма фундаментальным образом отличаются от всех соединений атомов, которые до настоящего времени являлись объектом экспериментальных и теоретических исследований физиков и химиков. Однако это различие, которое я считаю фундаментальным, может показаться малозначимым любому, кроме физика, сознающего, что законы химии и физики – сугубо статистические. Ведь именно со статистической точки зрения структура жизненно важных частей живых организмов столь отличается от любого кусочка материи, с которым мы, физики и химики, работаем физически в лабораториях или мысленно – за письменным столом[5]. Невозможно представить, что законы и закономерности, открытые подобным образом, могут непосредственно применяться к поведению систем, не обладающих структурой, на которой они основаны.
Не-физик вряд ли окажется способен хотя бы уловить – не говоря уже о том, чтобы оценить – различие в «статистической структуре», выраженное столь абстрактными терминами. Чтобы придать утверждению живость и цвет, позвольте мне упомянуть то, что позднее будет описано намного детальнее, а именно самую значимую составляющую живой клетки – хромосомную фибриллу, которую можно назвать апериодическим кристаллом. До настоящего времени в физике мы имели дело лишь с периодическими кристаллами. В сознании скромного физика это очень интересные и сложные объекты, они относятся к самым удивительным и хитроумным материальным структурам, которыми озадачила его неживая природа. Однако в сравнении с апериодическими кристаллами они незамысловаты и скучны. Различия в структурах можно сравнить с различием между обычными обоями, на которых один и тот же узор повторяется снова и снова с регулярной периодичностью, и искусной вышивкой, например гобеленом Рафаэля, где нет скучного повторения, а присутствует сложный, гармоничный, осмысленный рисунок, созданный великим мастером.
Называя периодические кристаллы одним из самых сложных объектов исследования, я имел в виду истинного физика. Органическая химия, исследуя все более замысловатые молекулы, намного ближе подобралась к тому «апериодическому кристаллу», который, на мой взгляд, является материальным носителем жизни. Неудивительно, что химики-органики уже внесли важный вклад в проблему жизни, в то время как физики не сделали почти ничего.
Теперь, обозначив вкратце главную идею, а точнее, пределы наших исследований, я опишу линию атаки. Я предлагаю вначале рассмотреть представления наивного физика об организмах – то есть идеи, что могут возникнуть в сознании физика, который, выучив свою физику, а точнее, статистическое основание науки, начинает размышлять о них и о том, как они себя ведут и функционируют, и в конце концов честно спрашивает себя, способен ли он посредством того, чему научился, с точки зрения своей относительно простой, четкой и скромной науки, внести какой-либо существенный вклад в данную проблему.
Оказывается, вполне способен. Далее нужно сравнить его теоретические ожидания с биологическими фактами. Выяснится, что, хотя в целом его идеи представляются весьма разумными, они нуждаются в значительной коррекции. Таким образом мы постепенно приблизимся к правильной точке зрения – точнее, если выразиться более скромно, точке зрения, которую я считаю правильной.
Я не уверен, что мой подход является самым лучшим и простым. Однако он мой. Я сам был «наивным физиком». И не смог отыскать более простого и ясного пути к цели, нежели моя кривая дорожка.
Хороший способ развить представления наивного физика – начать со странного, почти нелепого вопроса: почему атомы такие маленькие? Да, они действительно очень малы. Каждый фрагмент материи, с которым мы имеем дело в повседневной жизни, состоит из множества атомов. Чтобы донести этот факт до аудитории, подобраны многочисленные примеры, самый впечатляющий из которых принадлежит лорду Кельвину[6]. Представьте, что вы можете пометить молекулы в стакане воды; затем вылейте содержимое стакана в океан и тщательно перемешайте, чтобы равномерно распределить помеченные молекулы по семи морям. Если впоследствии вы наберете стакан воды в любом месте океана, то обнаружите в нем около сотни ваших меченых молекул. Разумеется, их будет не ровно 100 (даже если вычисления дают именно такой результат). Их будет 88, или 95, или 107, или 112, но вряд ли 50 или 150. Ожидаемое «отклонение», или «флуктуация», составит порядка корня квадратного из 100, то есть 10. Статистик выразит это так: вы обнаружите 100±10 молекул. Пока этот комментарий можно проигнорировать, однако позднее мы используем его в качестве иллюстрации статистического закона √n.
Реальный размер атомов[7] составляет примерно длину волны желтого света. Это сравнение существенно, поскольку длина волны грубо характеризует размеры мельчайшего объекта, видимого в микроскоп. Таким образом, подобный объект содержит тысячи миллионов атомов. Но почему атомы такие маленькие? Очевидно, данный вопрос является уловкой, поскольку в действительности он касается вовсе не размера атомов, а размера организмов, точнее, наших собственных тел. Атом мал в сравнении с «гражданской» единицей длины, например, ярдом или метром. В атомной физике обычно используют так называемый ангстрем (сокращенно Å), который составляет 10–10 метра, или, в десятичном представлении, 0,0000000001 метра. Диаметры атомов варьируют от 1 до 2 Å. «Гражданские» единицы, по сравнению с которыми атомы столь малы, тесно связаны с размерами наших тел. Согласно легенде, ярдом мы обязаны английскому королю-шутнику, которого советники спросили, какую единицу использовать. Он вытянул руку вбок и ответил: «Используйте расстояние от середины моей груди до кончиков пальцев, это подойдет». Правдива история или нет, но она важна для наших целей. Разумеется, король указал длину, сравнимую с его собственным телом, понимая, что любая другая будет неудобной. Несмотря на любовь к ангстремам, физик предпочитает слышать, что на его новый костюм потребуется шесть с половиной ярдов твида, а не шестьдесят пять тысяч миллионов ангстремов.
Таким образом, мы установили, что наш вопрос касается соотношения двух размеров – размера нашего тела и размера атома. С учетом неоспоримого главенства независимого существования атома, этот вопрос следует переформулировать так: почему наши тела столь велики в сравнении с атомом?
Представляю, как многие смышленые студенты-физики или химики оплакивали факт, что все наши органы чувств, составляющие вполне значимую часть организма, а следовательно с точки зрения вышеупомянутого соотношения состоящие из множества атомов, слишком грубы, чтобы ощутить влияние одиночного атома. Мы не можем увидеть, или почувствовать, или услышать отдельные атомы. Наши гипотезы на их счет заметно отличаются от непосредственных открытий, совершенных при помощи крупных органов чувств, и не могут быть проверены напрямую.
Обязательно ли это? Есть ли тому внутренняя причина? Можем ли мы проследить это состояние дел к некому первичному принципу, чтобы подтвердить и понять, почему ничто другое не совместимо с законами природы?
Наконец перед нами проблема, которую способен решить физик. Ответ на все эти вопросы утвердительный.
Если бы это было не так, если бы мы представляли собой организмы столь чувствительные, что один или несколько атомов сумели бы произвести ощутимое впечатление на наши чувства, – боже, какой была бы жизнь! Подчеркну: у подобного организма наверняка не развилось бы упорядоченное мышление, которое, пройдя немало ранних стадий, в конце концов сформировало бы, среди многих других идей, представление об атоме.
Мы выбираем именно этот момент, однако последующие рассуждения также применимы к работе других органов, а не только головного мозга и системы органов чувств. Тем не менее единственной действительно интересующей нас в нас самих вещью является то, что мы ощущаем, думаем и воспринимаем. По сравнению с физиологическим процессом, ответственным за мышление и чувства, остальные играют второстепенную роль, по крайней мере с точки зрения человека, если и не чисто объективной биологии. Более того, наша задача станет легче, если мы выберем для исследования процесс, тесно связанный с субъективными событиями, пусть и не осознавая истинной природы этого параллелизма. С моей точки зрения, он лежит за пределами естественных наук – и, вероятно, человеческого понимания.
Таким образом, перед нами возникает следующий вопрос: почему орган вроде нашего головного мозга, а также связанная с ним система органов чувств, должен состоять из невероятного числа атома, чтобы его физически изменчивое состояние соотносилось с высокоразвитым мышлением? Почему вышеупомянутая задача делает данный орган не совместимым с тем, чтобы являться – в целом или посредством периферических отделов, которые напрямую взаимодействуют с окружающей средой – инструментом достаточно тонким и чувствительным, чтобы зарегистрировать отдельный атом извне и отреагировать на него?
Причина такова: то, что мы называем мыслью, (1) само по себе упорядочено и (2) может использоваться лишь применительно к материалу, то есть восприятию или переживанию, обладающему определенным уровнем упорядоченности. Из этого вытекают два вывода. Во-первых, чтобы соотноситься с мышлением (как мой мозг соотносится с моей мыслью), физическая организация должна быть крайне упорядоченной, и это означает, что происходящие в ней события должны с высокой точностью подчиняться строгим физическим законам. Во-вторых, физические впечатления, которые производят на эту физически организованную систему внешние тела, очевидно, соотносятся с восприятием и опытом соответствующей мысли, формируя ее материал, как я выразился. Физические взаимодействия нашей системы с другими должны, как правило, сами обладать определенной степенью физической упорядоченности, то есть с определенной точностью подчиняться строгим физическим законам.
Почему же все это недостижимо для организма, состоящего из ограниченного числа атомов и способного ощутить воздействие одного или нескольких атомов?
Потому что мы знаем, что атомы постоянно находятся в неупорядоченном тепловом движении, которое, так сказать, противоречит упорядоченному поведению и не дает событиям, реализуемым небольшим числом атомов, соответствовать известным законам. Лишь в случае соединения невероятно большого числа атомов начинают действовать статистические законы, и они контролируют поведение этих скоплений с точностью, возрастающей вместе с ростом числа атомов. Именно таким образом события приобретают черты настоящей упорядоченности. Все физические и химические законы, играющие важную роль в жизни организмов, являются статистическими. Любой другой вид закономерности и упорядоченности нарушается и сводится на нет непрерывным тепловым движением атомов.
Позвольте проиллюстрировать это несколькими примерами, выбранными наугад из тысяч им подобных и потому, возможно, не самыми лучшими для читателя, который впервые слышит о таком положении вещей, – положении, столь же фундаментальном в современной физике и химии, как, например, клеточное строение организмов в биологии, или закон Ньютона в астрономии, или даже последовательность целых чисел – 1, 2, 3, 4, 5… – в математике. Следующие страницы вряд ли помогут новичку полностью понять и оценить предмет обсуждения, который связан с блистательными именами Уилларда Гиббса[8] и Людвига Больцмана[9] и обсуждается в учебниках в разделе «статистическая термодинамика».
Если заполнить вытянутую кварцевую трубку газообразным кислородом и поместить в магнитное поле, газ намагнитится. Я выбрал газ, поскольку случай с ним проще, чем с твердым веществом или жидкостью. Тот факт, что намагничивание в данном случае будет крайне слабым, не повлияет на теоретические рассуждения. Намагничивание происходит потому, что молекулы кислорода представляют собой маленькие магниты и ориентируются параллельно полю, как стрелка компаса. Но не думайте, будто они все выстраиваются параллельно. Удвоив силу поля, вы получите в вашем сосуде с кислородом двойную намагниченность, и она будет пропорционально возрастать по мере приближения к экстремально сильным полям.

Рис. 1. Парамагнетизм
Это наглядный пример чисто статистического закона. Ориентации, вызванной полем, постоянно противостоит тепловое движение, приводящее к произвольной ориентации. Результатом этой борьбы является незначительное преобладание острых углов между осью диполя и полем над тупыми углами. Ориентация отдельных атомов непрерывно меняется, однако в среднем, благодаря своему огромному количеству, они дают постоянное небольшое преобладание ориентации в направлении поля, пропорциональное этому полю. Этим блистательным объяснением мы обязаны французскому физику П. Ланжевену[10]. Проверить его можно следующим образом. Если наблюдаемая слабая намагниченность действительно является результатом противоборствующих явлений, а именно магнитного поля, желающего выстроить все молекулы параллельно, и теплового движения, которое стремится к произвольной ориентации, значит, можно усилить намагниченность, не повысив магнитное поле, а ослабив тепловое движение, то есть понизив температуру. Это подтверждает эксперимент, согласно ему намагниченность обратно пропорциональна абсолютной температуре, в количественном согласии с теорией (закон Кюри). Современное оборудование даже позволяет нам посредством понижения температуры настолько ослабить тепловое движение, что ориентирующее воздействие магнитного поля получит возможность если не проявить себя полностью, то достигнуть существенной доли «полной намагниченности». В данном случае мы уже не ждем, что удвоение силы поля удвоит намагниченность; последняя будет расти все меньше и меньше, приближаясь к так называемому насыщению. Это тоже подтверждает эксперимент.
Обратите внимание, что данное поведение полностью зависит от большого числа молекул, которые взаимодействуют, давая наблюдаемую намагниченность. В противном случае последняя не была бы постоянной, а весьма произвольно флуктуировала бы от секунды к секунде, свидетельствуя о переменных успехах в борьбе теплового движения и магнитного поля.
Заполнив нижнюю часть закрытого стеклянного сосуда туманом, состоящим из крошечных капель, вы увидите, что верхняя граница тумана будет постепенно опускаться с определенной скоростью, определяемой вязкостью воздуха и размером и удельной плотностью капель. Но изучив одну из капель под микроскопом, обнаружите, что она вовсе не опускается с постоянной скоростью, а совершает очень сложное движение, так называемое броуновское движение, которое лишь в среднем соотносится с общим оседанием.
Конечно, эти капли – не атомы, однако они достаточно малы и легки, чтобы ощутить влияние отдельных молекул, непрерывно бомбардирующих их поверхность. Поэтому капли отклоняются то в одну, то в другую сторону и лишь в среднем подчиняются действию силы тяжести.

Рис. 2. Оседающий туман

Рис. 3. Броуновское движение оседающей капли
Данный пример демонстрирует, какие забавные и хаотичные ощущения мы испытывали бы, если бы наши органы чувств воспринимали воздействие отдельных молекул. Существуют бактерии и другие организмы, которые столь малы, что на них этот феномен оказывает значительное влияние. Их движения определяются тепловыми капризами окружающей среды, у них просто нет выбора. Те из них, что обладают собственной способностью к перемещению, могут перебираться с места на место, однако с трудностями, поскольку тепловое движение швыряет их, словно утлую лодочку в бурном море.
На броуновское движение очень похоже явление диффузии. Представьте сосуд, заполненный водой, в которой растворено небольшое количество некоего окрашенного вещества, например перманганата калия, не в одинаковой концентрации, а как изображено на рис. 4, где точки обозначают молекулы растворенного вещества (перманганата) и концентрация падает слева направо. Если оставить этот сосуд в покое, начнется медленный процесс «диффузии», переносящий перманганат из левой части сосуда в правую, то есть из места с большей концентрацией в место с меньшей, пока вещество не распределится в воде равномерно.
Удивительной особенностью этого весьма простого и не слишком интересного процесса является то, что в его основе лежит не какая-то тенденция или сила, уводящая молекулы перманганата из более населенной области в менее населенную, словно жителей страны, переезжающих в свободные регионы. С нашими молекулами перманганата не происходит ничего подобного. Каждая ведет себя независимо от других, с которыми очень редко сталкивается. Каждая – как в населенной области, так и в пустой – постоянно испытывает удары молекул воды и постепенно движется в непредсказуемом направлении – иногда в область с большей концентрацией, порой в область с меньшей или вообще вбок. Перемещения такой молекулы часто сравнивают с движением на открытом пространстве слепого человека. Он одержим желанием «шагать», но не может выбрать направления, а потому непрерывно меняет свой курс.

Рис. 4. Диффузия слева направо в растворе с различной концентрацией
То, что это случайное блуждание всех без исключения молекул перманганата должно привести к регулярному потоку в направлении меньшей концентрации и – в конце концов – к равномерному распределению, на первый взгляд вызывает недоумение. Если поделить рис. 4 на тонкие срезы с приблизительно постоянной концентрацией, молекулы перманганата, содержащиеся в данном конкретном срезе в некий момент времени, за счет случайного движения с равной вероятностью переместятся влево или вправо. Однако благодаря этому плоскость, разделяющую соседние срезы, пересечет больше молекул, приходящих слева, нежели справа, – просто потому, что слева находится больше молекул, вовлеченных в случайное движение. И пока это соответствует действительности, результатом будет регулярный поток слева направо – до достижения равномерного распределения.
Если перевести эти рассуждения на язык математики, закон диффузии будет представлять собой дифференциальное уравнение с частными производными:
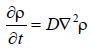
Я избавлю читателя от объяснений, хотя значение этого закона можно выразить простым языком. А именно: концентрация в любой конкретной точке возрастает или падает со временем пропорционально сравнительному избытку или недостатку концентрации в ее бесконечно малом окружении. Кстати, закон теплопроводности выглядит точно так же, только вместо концентрации стоит температура. Я привел этот суровый «математически строгий» закон, желая подчеркнуть, что его физическая точность должна, тем не менее, ставиться под сомнение в каждом конкретном случае. Он основан на случайности, и его правомерность приблизительна. Как правило, это очень хорошее приближение, но лишь благодаря огромному числу молекул, вовлеченных в явление. Чем меньше их количество, тем более сильных случайных отклонений следует ожидать – и они наблюдаются при неблагоприятных условиях.
Последний пример весьма похож на второй, однако представляет особый интерес. Легкое тело, подвешенное на длинной тонкой нити в равновесной ориентации, часто используется физиками для измерения слабых сил, которые отклоняют его от равновесия, электрических, магнитных или гравитационных сил, прикладываемых таким образом, чтобы повернуть тело вокруг вертикальной оси. Разумеется, выбор легкого тела должен соответствовать целям опыта. Непрерывные попытки повысить точность этих популярных «крутильных весов» выявили любопытный предел, интересный сам по себе. Если брать все более легкие тела и тонкие и длинные нити – чтобы равновесие было чувствительным к все более слабым силам, – предел достигается, как только подвешенное тело начинает ощущать влияние теплового движения молекул окружающей среды и исполнять непрерывный хаотический «танец» вокруг равновесного положения, подобно дрожащей капле. Подобное поведение не накладывает абсолютного предела на точность измерений, проведенных при помощи весов, однако подчеркивает практический предел. Неконтролируемое воздействие теплового движения конкурирует с воздействием измеряемой силы и делает отдельные наблюдаемые отклонения незначимыми. Следует провести множество измерений, чтобы исключить влияние броуновского движения на инструмент. Я считаю данный пример наиболее наглядным для нашего исследования, ведь наши органы чувств – тоже в определенном роде инструмент. Теперь мы видим, насколько бесполезными они станут, если обретут такую чувствительность.
Хочу добавить, что мог бы выбрать в качестве иллюстрации любой физический или химический закон из тех, что имеют значение для организма или его взаимодействий с окружающей средой. Подробное объяснение может оказаться более сложным, но суть будет той же, а потому описание станет монотонным.
Однако следует упомянуть одно важное численное утверждение относительно погрешности, которой следует ждать от любого физического закона, – правило √n. Вначале я проиллюстрирую его простым примером, а потом обобщу.
Если я предположу, что некий газ при определенных условиях – давлении и температуре – обладает определенной плотностью, и заявлю, что в определенном объеме (подходящем для какого-либо эксперимента) при этих условиях содержится n молекул газа, можете не сомневаться, что, проверив мое утверждение в некий момент времени, вы сочтете его ошибочным, с отклонением порядка √n. Соответственно, если n = 100, отклонение будет составлять около 10, а относительная ошибка – 10 %. Однако если n = 1 000 000, вы обнаружите отклонение около 1000, и относительная ошибка составит 0,1 %. Грубо говоря, данный статистический закон является весьма общим. Законы физики и физической химии неточны, и вероятная относительная ошибка для них составляет порядка  , где n есть число молекул, которые взаимодействуют, чтобы данный закон работал – и был справедливым в пространственных или временных (либо пространственно-временных) рамках, значимых для каких-либо рассуждений или эксперимента.
, где n есть число молекул, которые взаимодействуют, чтобы данный закон работал – и был справедливым в пространственных или временных (либо пространственно-временных) рамках, значимых для каких-либо рассуждений или эксперимента.
Из этого снова следует, что для того чтобы получать выгоду от достаточно точных законов, как во внутренних процессах, так и во взаимодействии с внешним миром, организм должен обладать крупной структурой. Иначе число взаимодействующих частиц будет слишком маленьким, а «законы» – неточными. Особенно строгим требованием является корень квадратный. Хотя миллион – весьма большое число, точность 1000 к 1 не кажется слишком высокой, если правило претендует на звание «закона природы».
Глава 2 Механизм наследственности
Оно [бытие] извечно; и законы Хранят, тверды и благосклонны Залоги дивных перемен.
Итак, мы пришли к выводу, что организм и испытываемые им биологически значимые процессы должны обладать в высшей степени «многоатомной» структурой и быть защищены от случайных «одноатомных событий». Это существенно для того, говорит нам «наивный физик», чтобы организм мог подчиняться достаточно точным физическим законам, на которых основана его удивительно регулярная и упорядоченная работа. Как эти выводы, достигнутые, биологически выражаясь, a priori, то есть с чисто физической точки зрения, согласуются с реальными биологическими фактами?
На первый взгляд кажется, будто данные выводы тривиальны. Лет тридцать назад какой-нибудь биолог вполне мог сказать, что хотя для популярного лектора уместно подчеркнуть значимость статистической физики, в частности для организма, сама идея банальна. Ведь не только тело взрослого представителя любого высшего вида, но и каждая составляющая его клетка содержит «космическое» число разнообразных атомов. И каждый наблюдаемый нами физиологический процесс внутри клетки или в ходе ее взаимодействия с окружающей средой, судя по всему – по крайней мере, так казалось тридцать лет назад, – затрагивает столь огромное число отдельных атомов и атомарных процессов, что соответствующим законам физики и физической химии ничто не грозит, несмотря на очень строгие требования, накладываемые статистической физикой на «большие числа». Эти требования я только что проиллюстрировал на примере правила √n.
Теперь мы знаем, что это мнение ошибочно. Как мы вскоре увидим, небольшие группы атомов – слишком небольшие, чтобы подчиняться точным статистическим законам, – играют важнейшую роль в упорядоченных и закономерных процессах в живом организме. Они управляют наблюдаемыми крупномасштабными особенностями, которые организм приобретает в ходе развития, определяют важные характеристики его работы – и во всем этом проявляются четкие и строгие биологические законы.
Следует начать с краткого описания ситуации, сложившейся в биологии, особенно в генетике. Иными словами, нужно суммировать нынешнее положение дел в предмете, которым я не владею. Тут ничего не поделаешь, и я прошу прощения, в первую очередь – у биологов, за дилетантский характер моей сводки. Кроме того, прошу дозволения изложить основные идеи в определенном смысле категорично. Нельзя ждать от бедного физика-теоретика компетентного обзора экспериментальных доказательств, которые включают, с одной стороны, множество длинных и красиво переплетающихся серий искусных экспериментов по скрещиванию, а с другой – прямые наблюдения за живой клеткой, выполненные с использованием изощренной современной микроскопии.
Позвольте мне использовать термин «структура» организма в том смысле, в каком биолог использует «четырехмерную структуру», имея в виду не только строение и работу этого организма во взрослом возрасте или на какой-то другой конкретной стадии, но его онтогенетическое развитие в целом, от оплодотворенной яйцеклетки до зрелости, когда он приступает к размножению. Известно, что всю эту четырехмерную структуру определяет строение одной-единственной клетки – оплодотворенной яйцеклетки. Более того, мы знаем, что структура, в сущности, определяется строением небольшой части этой клетки – ее ядра. В типичном «состоянии покоя» ядро обычно выглядит как сеть хроматина[11], распределенная по клетке. Однако в ходе жизненно важного процесса клеточного деления (митоз и мейоз) оно выглядит как набор частиц, обычно в форме фибрилл или палочек, называемых хромосомами. Их число составляет 8 или 12 – или, у человека, 48[12]. Однако мне следовало записать эти показательные числа как 2 (×) 4, 2 (×) 6… 2 (×) 24… и говорить о двух наборах, в привычном биологу смысле. Хотя отдельные хромосомы иногда отчетливо видны и различимы по форме и размеру, два их набора почти идентичны. Один набор приходит от матери (яйцеклетка), другой – от отца (оплодотворяющий сперматозоид). Именно эти хромосомы – или, возможно, лишь осевые скелетные фибриллы того, что мы видим под микроскопом как хромосомы, – содержат неким образом закодированную структуру будущего развития особи и ее функционирования в зрелом состоянии. Каждый полный набор хромосом содержит полный код, таким образом, две копии кода присутствуют в оплодотворенной яйцеклетке, которая является самой ранней стадией развития будущего организма.
Называя структуру хромосомных фибрилл кодом, мы имеем в виду, что придуманный Лапласом[13] всеведущий ум, мгновенно постигающий все причинно-следственные связи, может по ней узнать, вырастет ли из яйцеклетки при подходящих условиях черный петух или рябая курица, муха или кукуруза, рододендрон, жук, мышь или женщина. На вид все яйцеклетки удивительно схожи, а даже если и различаются – как в случае сравнительно крупных яиц птиц и рептилий, – эти различия затрагивают не основные структуры, а питательный материал, добавленный по очевидным причинам.
Разумеется, термин «код» является узким. Хромосомные структуры также участвуют в реализации развития, которое кодируют. Они объединяют в себе свод законов и исполнительную власть – или, если использовать другое сравнение, план архитектора и строительное мастерство.
Как хромосомы ведут себя во время онтогенеза?[14]
Рост организма осуществляется последовательными клеточными делениями. Такое клеточное деление называют митозом. В жизни клетки это не настолько частое событие, учитывая множество клеток, из которых состоит наше тело. Вначале рост осуществляется быстро. Яйцеклетка делится на две «дочерние клетки», они на следующих стадиях дают 4, затем 8, 16, 32, 64 и т. д. клеток. Частота делений неодинакова в различных частях растущего организма, и это нарушает регулярность числа клеток. Но на основании быстрого роста посредством простых вычислений можно установить, что в среднем 50–60 последовательных делений достаточно, чтобы получить число клеток, обнаруживаемое во взрослом человеке, или в десять раз больше, если учесть замену клеток в течение жизни. Таким образом, мое тело лишь на 50 или 60 поколений отстоит от яйцеклетки, которой я был.
Как ведут себя хромосомы в ходе митоза? Они дуплицируются – оба набора, обе копии кода. Данный процесс подробно изучен под микроскопом и очень важен, но слишком сложен, чтобы подробно описывать его здесь. Суть заключается в том, что каждая из двух «дочерних клеток» получает в наследство два полных набора хромосом, полностью идентичных набору родительской клетки. Поэтому все клетки тела имеют одинаковый хромосомный набор. Биолог простит мне удивительный случай мозаичности, который я опустил в своем кратком описании.
Хотя мы до сих пор плохо понимаем данный процесс, судя по всему, для работы организма важно, что каждая клетка, даже самая малозначимая, обладает полной (двойной) копией кода. Раньше в газетах писали, что во время своей африканской кампании генерал Монтгомери позаботился о том, чтобы каждый солдат его армии был подробно осведомлен о его планах. Если это соответствует действительности (а это вполне может ей соответствовать, учитывая сообразительность и надежность его отрядов), это отличная аналогия для нашего случая, в котором соответствующее утверждение в буквальном смысле верно. Самым удивительным фактом является двойственность хромосомного набора, поддерживаемая посредством митотических делений. Данную выдающуюся особенность генетического механизма особенно хорошо подчеркивает единственное исключение из правила, какое мы сейчас обсудим.
Вскоре после начала развития особи резервируется группа клеток, которая на более поздней стадии даст начало так называемым гаметам – сперматозоидам или яйцеклеткам, – необходимым для воспроизведения организма во взрослом возрасте. «Резервирование» означает, что пока они не выполняют никаких других функций и претерпевают значительно меньшее число митотических делений. Именно посредством исключительного, или редукционного, деления (мейоза) в зрелом возрасте из этих запасенных клеток образуются гаметы, обычно незадолго до того, как должна осуществиться сингамия. При мейозе двойной набор хромосом родительской клетки просто разделяется на два одинарных, по одному для каждой из двух дочерних клеток, или гамет. Иными словами, во время мейоза не происходит митотического удвоения числа хромосом, оно остается постоянным, и потому каждая гамета получает только половину – то есть одну полную копию кода, а не две (например, у человека только 24, а не 2 (х) 24 = 48).
Клетки с одним набором хромосом называют гаплоидными (от греч. ἁπλοῦς – одиночный). Таким образом, гаметы гаплоидны, а обычные клетки тела диплоидны (от греч. διπλοῦς – двойной). Иногда встречаются особи с тремя, четырьмя… в общем, с множественными наборами хромосом. Их называют триплоидными, тетраплоидными… полиплоидными.
В ходе сингамии гаплоидная мужская гамета (сперматозоид) и гаплоидная женская гамета (яйцеклетка) сливаются в оплодотворенную диплоидную яйцеклетку. Она получает один хромосомный набор от матери и один – от отца.
Еще один момент требует уточнения. Для нас он не слишком важен, однако представляет значительный интерес, поскольку демонстрирует, что практически полный код «структуры» содержится в каждом одинарном наборе хромосом.

Рис. 5. Чередование поколений
В некоторых случаях непосредственно после мейоза не происходит оплодотворения, и гаплоидная клетка («гамета») претерпевает многочисленные митотические деления, приводящие к формированию полностью гаплоидной особи. Например, самец пчелы – трутень – развивается партеногенетически, то есть из неоплодотворенных, а значит, гаплоидных яйцеклеток матки. У трутня нет отца! Все клетки его тела гаплоидны. Можно назвать его огромным сперматозоидом – и действительно, как известно, это его единственная функция. Однако данная точка зрения нелепа. Такой случай не уникален. Существуют семейства растений, у которых гаплоидная гамета, образующаяся путем мейоза и называемая спорой, падает на землю и, подобно семени, развивается в истинно гаплоидное растение, по размерам сравнимое с диплоидным. На рис. 5 приведено схематическое изображение мха, распространенного в наших лесах. Облиственная нижняя часть представляет собой гаплоидное растение, называемое гаметофитом, потому что на его верхнем конце развиваются половые органы и гаметы, которые путем взаимного оплодотворения дают диплоидное растение – голый стебель со споровой коробочкой наверху. Когда коробочка раскрывается, споры попадают на землю, из них развивается облиственный стебель и т. д. Эту последовательность событий называют чередованием поколений. Можно взглянуть подобным образом на человека и животных. Однако «гаметофит» у них, как правило, представляет собой короткоживущее одноклеточное поколение, сперматозоид или яйцеклетку. Наше тело соответствует спорофиту. Наши «споры» – запасенные клетки, из них посредством мейоза образуется одноклеточное поколение.
Важным, действительно судьбоносным событием в процессе размножения особи является не оплодотворение, а мейоз. Один набор хромосом достается от отца, другой – от матери. Ни случай, ни рок не может повлиять на это. Каждый человек[15] обязан половиной своего наследственного материала отцу и половиной – матери. То, что обычно одна из них доминирует, объясняется другими причинами, до которых мы еще дойдем. Разумеется, самый простой пример такого доминирования – пол.
Но если проследить вашу наследственность до бабушки и дедушки, ситуация изменится. Возьмем набор хромосом моего отца, точнее, одну их пару, например № 5. Это верная копия № 5, которую отец получил от своего отца, или № 5, какую отец получил от своей матери. Вопрос был решен с вероятностью 50:50 в ходе мейоза, случившегося в организме моего отца в ноябре 1886 года и произведшего сперматозоид, который несколько дней спустя принял участие в зачатии меня. Аналогичную же историю можно рассказать про хромосомы № 1, 2, 3… 24 из отцовского набора и, mutatis mutandis[16], из материнского набора. Более того, все 48 случаев независимы друг от друга. Даже если бы я знал, что отцовская хромосома № 5 попала ко мне от моего деда Йозефа Шредингера, хромосома № 7 имеет равные шансы также принадлежать ему – или его жене Марии, в девичестве Богнер.
Однако чистая случайность получила еще больше возможностей смешать в потомке наследие дедов и бабок, чем может показаться на основании вышеизложенного. Ранее мы полагали, будто конкретная хромосома целиком досталась нам от деда или от бабки, иными словами, что отдельные хромосомы передаются неделимыми. В действительности это не так, по крайней мере, не всегда. Прежде чем разойтись в ходе редукционного деления, например, происходящего в организме отца, любые две «гомологичные» хромосомы вступают в близкий контакт друг с другом и обмениваются целыми участками, как показано на рис. 6. Благодаря этому процессу, называемому кроссинговер, два признака, расположенные в соответствующих частях данной хромосомы, окажутся разделены у внука, который получит одно из них от деда, а другое от бабки. Процесс кроссинговера снабдил нас важной информацией о расположении признаков на хромосомах. Для полноценного рассмотрения следовало бы очертить концепции, которые будут представлены в следующей главе (такие как гетерозиготность, доминантность и т. д.), однако это выведет нас за рамки данной книги, а потому позвольте сразу перейти к сути.

Рис. 6. Кроссинговер. Слева: контакт между двумя гомологичными хромосомами. Справа: гомологичные хромосомы после обмена и расхождения
Если бы не кроссинговер, два признака, за которые отвечает одна и та же хромосома, всегда передавались бы вместе, ни один потомок не смог бы получить один из них, не унаследовав при этом другой. Однако два признака, находящиеся на различных хромосомах, будут разделены с вероятностью 50:50 – или со 100 % вероятностью, если расположены на гомологичных хромосомах одного предка, которые не могут передаться вместе.
Кроссинговер нарушает эти правила. Таким образом, вероятность данного события можно оценить, тщательно отслеживая процентный состав потомства в длительных экспериментах по скрещиванию. Анализируя статистику, следует принять рабочую гипотезу, что «связь» между двумя признаками, расположенными на одной хромосоме, нарушается тем реже, чем ближе они находятся друг к другу. В таком случае ниже вероятность, что точка обмена окажется между ними, в то время как признаки, расположенные на противоположных концах хромосом, разделяются при каждом кроссинговере. То же самое верно для рекомбинации признаков, расположенных на гомологичных хромосомах одного предка. Таким образом, на основании «статистики связи» можно получить некую «карту признаков» каждой хромосомы.
Эти ожидания полностью подтвердились. В проведенных экспериментах, в основном – но не только – на дрозофиле, изученные признаки действительно разделяются на группы, не сцепленные друг с другом, число которых равняется числу отдельных хромосом (четыре у дрозофилы). В каждой группе можно составить линейную карту признаков, количественно соответствующую степени сцепления между любыми двумя признаками в данной группе. Поэтому нет сомнений, что они действительно расположены на хромосоме, причем линейно, что предполагает ее палочковидная форма.
Разумеется, изображенный здесь схематический механизм наследственности выглядит поверхностным и бесцветным, даже слегка наивным. Ведь мы не сказали, что именно называем «признаком». Кажется неправильным и невозможным расчленять на отдельные «признаки» структуру организма, который по существу является единым, «целым». В любом конкретном случае мы утверждаем, что пара предков различалась по явно заданным характеристикам (например, у одного были голубые глаза, а у другого – карие), и потомок в данном смысле похож на одного из них. То, что мы обнаруживаем в хромосоме, и есть причина этого различия. (Технический термин для этого – «локус», или, если имеется в виду лежащая в его основе гипотетическая структура, «ген».) На мой взгляд, различие в признаках само по себе является фундаментальной концепцией, а не признаком, вопреки очевидному лингвистическому и логическому противоречию данного утверждения. Различия в признаках дискретны, как мы поймем из следующей главы, когда будем обсуждать мутации, и я надеюсь, что приведенная здесь сухая схема обретет жизнь и заиграет красками.
Мы только что ввели термин «ген» – гипотетический материальный носитель определенной наследственной характеристики. Теперь следует подчеркнуть два момента, в высшей степени значимые для нашего исследования. Во-первых, размер – а точнее, максимальный размер – такого носителя; иными словами, до сколь малого объема мы сможем проследить местоположение признака? Во-вторых, постоянство гена, которое следует из устойчивости наследственной структуры.
Что касается размера, то существуют две независимые оценки. Одна опирается на генетические доказательства (эксперименты по скрещиванию), а другая – на цитологические свидетельства (прямое микроскопическое наблюдение). Первая по сути своей несложна. После того как мы выявим указанным выше способом на конкретной хромосоме достаточное число различных («широкомасштабных») признаков (например, у мушки-дрозофилы), останется разделить измеренную длину этой хромосомы на число признаков и умножить на поперечное сечение. Разумеется, мы считаем различными только те признаки, которые время от времени разделяет кроссинговер, а потому они не могут определяться одной и той же структурой – микроскопической или молекулярной. С другой стороны, очевидно, что наша оценка может дать лишь максимальный размер, поскольку число признаков, обнаруженных генетическим анализом, постоянно растет.
Другая оценка, пусть и основанная на микроскопическом наблюдении, в действительности является косвенной. Определенные клетки дрозофилы, а именно клетки слюнных желез, по какой-то причине сильно увеличены, как и их хромосомы. На этих фибриллах можно различить плотный рисунок из поперечных темных полос. С. Д. Дарлингтон[17] отметил, что число этих полос (в его случае – 2000) хотя и намного превышает число генов, расположенных на хромосоме, как установлено в экспериментах со скрещиванием, но находится в пределах того же порядка. Он склонен считать эти полосы настоящими генами (или границами между ними). Поделив длину хромосомы, измеренную в клетке нормального размера, на число полос (2000), он получает объем гена, равный кубу с гранью 300 Å. Учитывая приблизительность оценки, можно предположить, что такой же результат был получен и первым способом.
Подробное обсуждение связи статистической физики со всеми перечисленными фактами – а точнее, связи этих фактов с использованием статистической физики в живой клетке – последует позднее. Но позвольте сейчас привлечь внимание к тому, что 300 Å – лишь 100–150 атомных расстояний в жидкости или твердой материи, а значит, ген определенно не может содержать больше миллиона или нескольких миллионов атомов. Это число слишком мало (с точки зрения √n), чтобы обеспечить упорядоченное поведение в соответствии со статистической физикой, а следовательно, и физикой вообще. Оно слишком мало, даже если все эти атомы играют одну и ту же роль, как в газе или капле жидкости. Но ген – определенно не гомогенная капля жидкости. Это крупная белковая молекула, в которой каждый атом, каждый остаток, каждое гетероциклическое кольцо выполняет особую функцию, отличную от функций других схожих атомов, остатков или колец. Так считают ведущие генетики, например Холдейн и Дарлингтон, и вскоре нам придется обратиться к генетическим экспериментам, практически доказывающим эту точку зрения.
Теперь второй важный вопрос: какой степени постоянства ожидать от наследуемых признаков, а значит, и материальных структур, которые их переносят?
Ответ можно дать без специальных исследований. Факт, что мы обсуждаем наследуемые признаки, свидетельствует о признании их почти незыблемого постоянства. Не надо забывать, что ребенку передается от родителя не просто та или иная черта – кривой нос, короткие пальцы, предрасположенность к ревматизму, гемофилии, дихромазии и т. п. Подобные признаки мы можем выбрать для изучения законов наследственности. Однако в действительности речь идет о полной (четырехмерной) структуре «фенотипа», видимой и проявляемой природе особи, какая воспроизводится без значимых изменений от поколения к поколению, неизменная на протяжении веков – хотя и не десятков тысячелетий, – всякий раз переносимая материальной структурой ядер двух клеток, которые объединяются, чтобы дать оплодотворенную яйцеклетку. Это чудо – уступающее лишь другому чуду, оно, пусть и тесно связанное с первым, лежит в иной плоскости. Я имею в виду тот факт, что мы, чье существование основано на чудесных взаимодействиях такого рода, все же обладаем способностью в значительной степени постигать их. Думаю, это понимание – первого чуда – может стать почти полным. Однако второе может остаться непостижимым для человека.
Глава 3 Мутации
Но эти превращенья в свой черед немеркнущими мыслями украсьте.
Изложенные нами общие факты в поддержку стойкости структуры генов, вероятно, слишком привычны, чтобы казаться удивительными либо убедительными. В данной ситуации верна пословица: исключение подтверждает правило. Если бы дети всегда были похожи на родителей, мы бы лишились не только красивых экспериментов, раскрывших нам подробный механизм наследственности, но и великого колоссального эксперимента природы, которая создает виды путем естественного отбора и выживания наиболее приспособленных.
Давайте используем последний важный вопрос в качестве отправной точки для представления значимых фактов – вновь прошу прощения и напоминаю, что я не биолог.
Мы точно знаем, что Дарвин ошибался, считая незначительные, постоянные, случайные вариации, имеющие место даже в самой гомогенной популяции, материалом естественного отбора. Доказано, что они не наследуются. Это важный факт, достойный краткого примера. Если взять урожай чистого сорта ячменя, измерить на всех колосьях длину ости и представить результаты измерений на графике, то получится колоколообразная кривая, как показано на рис. 7, где число колосьев соответствует определенной длине ости. Иными словами, преобладает определенная средняя длина, отклонения от которой в какую-либо сторону происходят с некой частотой.

Рис. 7. Статистика длин остей у чистой линии ячменя. Группа, выбранная для посева, показана черным цветом (Это не результаты некоего эксперимента, а теоретический пример)
Теперь выберем группу колосьев (показана черным цветом), чьи ости заметно длиннее среднего и чьего количества достаточно, чтобы высадить их в поле и получить новый урожай. Проделав аналогичные измерения, Дарвин ожидал бы, что кривая сместится вправо. Он ожидал бы путем селекции добиться увеличения средней длины ости. При использовании чистой линии ячменя результат будет другим. Новая статистическая кривая окажется идентичной первой. То же самое повторится, если выбрать колосья с особенно короткой остью. Отбор не оказывает влияния, поскольку незначительные, постоянные вариации не наследуются. Они не определяются структурой наследственного материала и происходят случайно. Однако около сорока лет назад де Фриз[18] обнаружил, что даже в потомстве чистых линий появляется очень небольшое число особей – две-три на десятки тысяч – с малыми, но «скачкообразными» изменениями. Слово «скачкообразный» характеризует не величину изменения, а дискретность, поскольку не обнаруживается промежуточных форм между исходным и измененным состоянием. Де Фриз назвал это мутацией. Дискретность является важным фактом. Она напоминает физику квантовую теорию: между соседними энергетическими уровнями нет промежуточных энергий. Физик метафорически назвал бы мутационную теорию де Фриза «квантовой теорией биологии». Позднее мы увидим, что это определение вовсе не столь метафорично. Мутации действительно происходят за счет квантовых скачков в молекуле гена. Однако квантовой теории исполнилось всего два года, когда в 1902 году де Фриз опубликовал свое открытие. Неудивительно, что лишь следующее поколение ученых обнаружило тесную связь между ними!
Мутации наследуются так же, как исходные, неизмененные признаки. Например, в рассмотренном выше первом урожае ячменя может появиться несколько колосьев с остями, заметно выходящими за рамки изменчивости, показанные на рис. 7, вообще без ости. Они могут быть носителями мутации де Фриза, которая передастся потомству, а значит, все их потомки будут лишены ости.
Таким образом, мутация представляет собой изменение наследия и должна определяться неким изменением наследственного материала. На самом деле, большинство важных экспериментов по скрещиванию, объяснивших нам механизм наследственности, заключались в анализе потомства, полученного скрещиванием согласно разработанной схеме, носителей мутации (во многих случаях – нескольких мутаций) с немутантными особями или с носителями других мутаций. Кроме того, поскольку мутации наследуются, они являются подходящим материалом, с которым может работать естественный отбор, производя новые виды, как описал Дарвин, путем вымирания неприспособленных и выживания сильнейших. Нужно лишь заменить в теории Дарвина «незначительные случайные вариации» на «мутации» (так же, как квантовая теория заменяет «постоянный перенос энергии» «квантовым переходом»). Во всех прочих аспектах теория Дарвина нуждалась в незначительной корректировке, если я правильно понимаю точку зрения большинства биологов. Активно обсуждался вопрос, способствует ли естественному отбору (или даже доминирует ли над ним) явная склонность мутаций происходить в благоприятном направлении. У меня на данный момент нет своей точки зрения, однако следует отметить, что возможность «направленности» мутаций далее не учитывается. Более того, я не могу уделить внимание взаимодействию генов-«переключателей» и «полигенов», как бы ни были они важны для механизма естественного отбора и эволюции.
Теперь следует рассмотреть фундаментальные факты и понятия, касающиеся мутаций, вновь в немного категоричной форме, не затрагивая напрямую подтверждающие их экспериментальные доказательства.

Рис. 8. Гетерозиготный мутант. Крестиком показан ген с мутацией
Разумно предположить, что некую наблюдаемую мутацию вызывает изменение определенного участка одной из хромосом. Так и есть. Важно отметить, что мы точно знаем: это изменение затрагивает лишь одну хромосому, но не соответствующий «локус» на гомологичной хромосоме. Это схематично показано на рис. 8; мутировавший локус обозначен крестиком. Факт, что мутация происходит только в одной хромосоме, можно доказать, скрестив мутантную особь («мутанта») с нормальной. Половина потомства будет обладать мутантным признаком, а другая половина – нормальным. Этого следует ожидать вследствие расхождения двух хромосом у мутанта во время мейоза, как схематично показано на рис. 9. Это «фамильное дерево», на котором каждая особь (из трех последовательных поколений) представлена парой рассматриваемых хромосом. Имейте в виду, что если у мутанта затронуты обе хромосомы, все его потомки получат одинаковое (смешанное) наследие, отличное от родительского.

Рис. 9. Наследование мутации. Поперечные прямые линии показывают передачу хромосомы, двойные линии – передачу мутантной хромосомы. Неучтенные хромосомы третьего поколения пришли от партнеров второго поколения, не включенных в схему. Предполагается, что это не родственники, лишенные мутаций
Однако эксперименты в данной области не столь просты, как может показаться. Их затрудняет второй важный фактор, а именно то, что мутации очень часто латентны. Что это означает?
У мутанта две «копии кода» уже не идентичны, а представляют собой два различных «прочтения», или «версии», по крайней мере в одном конкретном месте. Следует отметить, что, как бы заманчиво это ни выглядело, нельзя считать исходную версию «ортодоксальной», а мутантную – «еретической». В принципе мы должны полагать, что они имеют равное право на существование, поскольку нормальные признаки также возникли благодаря мутациям.

Рис. 10. Гомозиготный мутант, представленный одной четвертью потомства, появившегося путем самооплодотворения гетерозиготного мутанта (рис. 8) либо скрещивания двух гетерозиготных мутантов
В действительности «структура» особи, как правило, определяется той или иной версией, которая может быть нормальной либо мутантной. Определяющую версию называют доминантной, другую – рецессивной. Иными словами, мутация является доминантной или рецессивной в зависимости от того, меняет ли она непосредственно структуру или нет.
Рецессивные мутации встречаются намного чаще доминантных и крайне важны, хотя поначалу никак не проявляют себя. Чтобы повлиять на структуру, они должны присутствовать на обеих хромосомах (см. рис. 10). Подобные особи появляются, когда друг с другом скрещиваются два одинаковых рецессивных мутанта или когда мутант скрещивается сам с собой. Такое бывает у гермафродитных растений и даже случается спонтанно. В данном случае около одной четверти потомков будут носителями двух мутантных хромосом, а значит, у них проявится мутантный признак.
Полагаю, для ясности следует объяснить несколько технических терминов. То, что я назвал версией кода – исходной или мутантной, – называют аллелью. Если версии различаются, как на рис. 8, особь называют гетерозиготной по данному локусу. Если версии одинаковы, как у немутантной особи или в случае, показанном на рис. 10, такие организмы называют гомозиготными. Так, рецессивная аллель влияет на структуру только в гомозиготном состоянии, в то время как доминантная аллель дает одну и ту же структуру и в гомозиготном, и в гетерозиготном состоянии.
Цвет часто доминирует над бесцветностью (или белизной). Например, горох обладает белыми цветками, только когда содержит в обеих соответствующих хромосомах «рецессивную аллель, отвечающую за белизну», то есть когда он «гомозиготен по белизне». Этот признак будет наследоваться, и все потомки данного растения станут цвести белым цветом. Однако присутствие одной «красной аллели» (в то время как другая остается белой – у «гетерозиготы») – равно как и две «красных аллели» (у «гомозиготы») – заставит горох цвести красными цветками. Различие последних двух случаев проявится только в потомстве, когда красные гетерозиготы дадут несколько белых потомков, а красные гомозиготы сохранят признак.
Факт, что две особи могут обладать совершенно одинаковой внешностью, но разной наследственностью, является столь важным, что следует точно разграничить эти понятия. Генетики говорят, что у таких особей одинаковый фенотип, однако разный генотип. Таким образом, вышеизложенное можно суммировать кратким, но в высшей степени техническим утверждением: рецессивная аллель влияет на фенотип лишь в том случае, когда генотип гомозиготен.
Мы будем использовать эти технические термины, однако при необходимости будем напоминать читателю их значение.
В гетерозиготном состоянии рецессивные мутации – неподходящий материал для естественного отбора. Если они вредны, как часто и бывает, то все равно сохраняются в силу своей латентности. Поэтому значительное количество неблагоприятных мутаций может накапливаться, не причиняя непосредственного вреда. Однако они передаются половине потомков – и это имеет большое значение для человека, домашнего скота, птицы и любого другого вида, физическое состояние которого нас заботит. На рис. 9 предполагается, что особь мужского пола, например я сам, несет рецессивную вредную мутацию в гетерозиготном состоянии, то есть она не проявляется. Предположим, у моей жены нет такой мутации. Тогда половина наших детей (второе поколение) окажется носителями мутации – снова гетерозиготной. Если все они опять выберут себе партнеров без мутаций (не показаны на схеме для ясности), около четверти наших внуков будут обладать гетерозиготной мутацией.
Никакого вреда не случится, если только особи с одинаковой мутацией не скрестятся друг с другом. В таком случае, как показывают нехитрые расчеты, четверть их детей будет гомозиготной, и у них неблагоприятная мутация проявится. Помимо самооплодотворения, которое возможно лишь у растений-гермафродитов, наибольшую опасность будет представлять брак между моим сыном и моей дочерью. Каждый из них обладает равными шансами оказаться или не оказаться носителем опасной мутации, и четверть таких кровосмесительных союзов приведет к тому, что у четверти их потомков она проявится. Таким образом, вероятность заболеть у родившегося от подобного союза ребенка окажется 1:16.
Аналогичным образом вероятность заболеть будет составлять 1:64 у потомка союза между двумя моими («чистокровными») внуками, являющимися двоюродными братом и сестрой. Это не слишком высокая вероятность, и второй вариант обычно считают приемлемым. Но не забывайте, что мы рассмотрели последствия лишь одного латентного повреждения у одного партнера предковой пары («я и моя жена»). В действительности они оба скорее всего несут несколько подобных латентных изъянов. Если вы точно знаете, что являетесь обладателем некой мутации, не забывайте, что 1 из 8 ваших кузенов и кузин также может нести ее! Эксперименты на растениях и животных свидетельствуют о том, что, помимо сравнительно редких серьезных пороков, существует множество не столь значительных дефектов, которые в совокупности способны повредить потомку близкородственного скрещивания. Поскольку мы больше не избавляемся от неудачных экземпляров, как это делали суровые спартанцы в горах Тайгет, следует относиться к этим вопросам серьезно, в случае человека, для которого не всегда соблюдается выживание сильнейших, а то и реализуется противоположная ситуация. Противодействующее отбору влияние современного массового убийства здоровых молодых людей всех национальностей перевешивает рассуждения о том, что в более примитивных условиях война могла производить положительный эффект, способствуя выживанию наиболее приспособленного клана.
Удивителен факт, что рецессивная аллель в гетерозиготном состоянии полностью подавляется доминантной и не дает видимого эффекта. Следует отметить, что из этого правила есть исключения. Если скрестить гомозиготный белый львиный зев с гомозиготным малиновым, все потомки первого поколения будут обладать «промежуточным» цветом, то есть розовым, а не малиновым, как можно было бы ожидать. Намного более важным примером одновременного проявления двух аллелей являются группы крови, но мы не будем здесь на них останавливаться. Не удивлюсь, если в конце концов выяснится, что рецессивность обладает уровнями и ее проявления зависят от чувствительности тестов, которые мы используем для анализа «фенотипа».
Уместно вспомнить раннюю историю генетики. Суть теории, закон наследования последующими поколениями признаков, по которым различаются родители, и – что особенно важно – разграничение рецессивных и доминантных признаков сформулировал всемирно известный аббат-августинец Грегор Мендель (1822–1884). Он ничего не знал о мутациях и хромосомах. В своем монастырском саду в Брюнне (Брно) Мендель проводил эксперименты на горохе: сажал различные разновидности, скрещивал и наблюдал за потомками в 1, 2, 3-м… поколениях. Можно сказать, он экспериментировал с мутантами, предоставленными ему природой. В 1886 году Мендель опубликовал результаты в «Трудах Брюннского общества естествоиспытателей». Никто не обращал особого внимания на хобби аббата – и уж точно никто и представить не мог, что в XX веке его открытие станет путеводной звездой для совершенно нового научного направления, одного из самых интересных в наши дни. Его статью забыли. Лишь в 1900 году о ней одновременно вспомнили Корренс[19] (Берлин), де Фриз (Амстердам) и Чермак[20] (Вена).
До настоящего момента мы в основном говорили о вредоносных мутациях, которые, возможно, являются самыми многочисленными; однако следует отметить, что встречаются и полезные мутации. Если спонтанная мутация есть небольшой шажок в развитии вида, то складывается впечатление, что определенные изменения «подбираются» весьма произвольным образом, рискуя причинить вред, в случае чего они тут же уничтожаются. Из этого следует важный вывод. Чтобы являться подходящим материалом для естественного отбора, мутации должны происходить редко – что и наблюдается в действительности. Если бы они случались так часто, что существовала бы вероятность появления у одной особи, скажем, десятка мутаций, вредоносные, как правило, преобладали бы над полезными, и вид не улучшался бы в результате отбора, а оставался бы прежним или вымирал бы. Умеренный консерватизм, являющийся следствием высокой стабильности генов, играет существенную роль. Можно провести аналогию с работой крупной производственной установки на фабрике. Чтобы выработать лучшие методы, необходимо применять инновации, в том числе неопробованные. Но чтобы установить, улучшит или ухудшит инновация работу, нужно пробовать их по одной, не изменяя другие детали механизма.
Теперь рассмотрим блистательный цикл генетических исследований, которые внесут наиболее значимый вклад в наш анализ.
Процент мутаций у потомства, так называемую частоту мутаций, можно повысить в сравнении с низкой природной частотой мутаций, облучив родителей рентгеновскими или гамма-лучами. Полученные таким образом мутации ничем (кроме количества) не отличаются от спонтанных, и складывается впечатление, что рентгеновские лучи также могут вызывать «природные» мутации. У дрозофилы многие особые мутации спонтанно происходят снова и снова в больших культурах; их местоположение картировали на хромосоме и дали им специальные названия. Более того, были обнаружены «множественные аллели», то есть две и более различных «версий» и «прочтений» – помимо нормальной, немутантной – одного участка хромосомного кода. Это означает существование не двух, но трех и более вариантов данного конкретного «локуса», любые два из которых относятся друг к другу как «доминантные/рецессивные», когда одновременно встречаются в соответствующих локусах двух гомологичных хромосом.
Эксперименты с мутациями, индуцированными рентгеновским излучением, создают впечатление, что каждый «переход», скажем, от нормальной особи к некому мутанту или наоборот, обладает своим «рентгеновским коэффициентом», выражающим процент потомков, которые мутировали данным образом при воздействии на родителей стандартной дозы рентгеновского излучения до зачатия.
Более того, законы, управляющие частотой индуцированных мутаций, просты и наглядны. Здесь я буду придерживаться сообщения Н.В Тимофеева-Ресовского[21], опубликованного в «Биологических обзорах», т. IX за 1934 год. В значительной степени оно касается блистательной работы самого автора. Первый закон таков:
(1) Рост частоты мутаций точно пропорционален дозе облучения, поэтому можно говорить [как это сделал я] о коэффициенте роста.
Мы настолько привыкли к простой пропорциональности, что можем недооценить далеко идущие следствия этого нехитрого закона. Чтобы их понять, давайте вспомним, что, например, цена товара отнюдь не всегда пропорциональна его количеству. При обычных обстоятельствах продавец настолько удивится, если вы купите у него шесть апельсинов, что, когда вы решите взять целую дюжину, может отдать ее дешевле двойной цены за шесть штук. Во времена дефицита происходит обратное. В настоящем случае мы делаем вывод, что первая половинная доза радиации, которая, скажем, вызывает мутации у одного из тысячи потомков, не затрагивает всех остальных – не делает их более склонными или устойчивыми к мутации. Иначе повторная половинная доза не вызвала бы снова лишь одну мутацию на тысячу. Таким образом, мутация не является кумулятивным эффектом, к которому приводят последовательные небольшие дозы радиации, усиливающие друг друга. Она должна представлять собой единичное событие, происходящее в одной хромосоме во время облучения. Что это за событие?
На данный вопрос отвечает второй закон, а именно:
(2) Если варьировать характеристики (длину волны) излучения в широких пределах, от мягких рентгеновских лучей до весьма жестких гамма-лучей, коэффициент останется неизменным, при условии, что вы будете использовать одну и ту же дозу в так называемых рентгенах. То есть если определите дозу, измерив общее количество ионов, производимых на единицу объема в соответствующем стандартном веществе в том же месте и в то же время, когда подвергнутся облучению родители.
В качестве стандартного вещества используют воздух – не только ради удобства, но и потому, что органические ткани построены из элементов с таким же атомным весом. Нижний[22] порог величины ионизации или сопряженных процессов (возбуждений) в тканях получают, умножив уровень ионизации воздуха на отношение плотностей. Таким образом, очевидно – и более серьезное исследование подтверждает это, – что единичное событие, вызывающее мутацию, представляет собой ионизацию (или сходный процесс), происходящую в некоем «критическом» объеме половой клетки. Каков этот критический объем? Его можно оценить на основании частоты мутаций посредством следующего рассуждения: если доза в 50 000 ионов на 1 см3 приводит к вероятности возникновения мутации 1:1000 в любой конкретной гамете (что оказалась в области облучения), «цель», в которую следует «попасть» ионизации для появления этой мутации, занимает лишь одну пятидесятимиллионную долю сантиметра кубического. Эти числа не соответствуют действительности и приведены исключительно ради примера. Реальную оценку дает М. Дельбрюк[23] в статье Дельбрюка, Н. В. Тимофеева-Ресовского и К. Г. Циммера[24], которая также является основным источником теории, изложенной в следующих двух главах. Он приводит значение десять средних атомных расстояний в кубе, где содержится всего лишь около 103 – тысяча – атомов. Простейшая интерпретация данного результата заключается в том, что существует высокая вероятность вызвать мутацию, если ионизация (или возбуждение) имеет место «не далее чем в десяти атомах» от данного конкретного места хромосомы. Обсудим это подробнее.
В отчете Тимофеева-Ресовского есть практический намек, который я обязан упомянуть, пусть он и не имеет отношения к нашему текущему исследованию. В реальной жизни человек часто подвергается рентгеновскому излучению. Сопряженные с этим прямые опасности – такие как ожоги, лучевой рак, бесплодие – хорошо известны, для защиты от них используют свинцовые экраны, фартуки и т. п., особенно медсестры и врачи, вынужденные регулярно иметь дело с излучением. Суть в том, что, несмотря на успешную защиту от непосредственных угроз, также существует непрямая опасность возникновения вредоносных мутаций в половых клетках – подобных тем, что мы обсуждали, рассматривая неблагоприятный исход близкородственного скрещивания. Выражусь резко, пусть и немного наивно: опасность брака между двоюродными братьями и сестрами может возрасти в том случае, если их бабушка долгое время работала медсестрой и занималась рентгеном. Не следует принимать это близко к сердцу. Однако обществу нужно тревожиться о любой возможности постепенно поразить человеческую расу нежелательными латентными мутациями.
Глава 4 Квантово-механическое доказательство
И пусть твой дух как пламя вознесен, подобьями довольствуется он.
Итак, с помощью чудесного тонкого инструмента – рентгеновских лучей, благодаря которым, как помнят физики, тридцать лет назад была детально изучена атомная решетка кристаллов, биологи и физики, объединив усилия, опустили верхний предел размера микроскопических структур, отвечающих за определенные крупномасштабные признаки особи – «размер гена». Теперь перед нами встал серьезный вопрос: как с точки зрения статистической физики примирить те факты, что структура генов включает сравнительно небольшое число атомов (порядка 1000, а то и меньше), но тем не менее проявляет в высшей степени регулярную и упорядоченную активность, с постоянством и стабильностью, граничащими с чудом?
Позвольте мне еще раз подчеркнуть эту удивительную ситуацию. Несколько представителей династии Габсбургов обладают странной деформацией нижней губы – «габсбургской губой». Под покровительством императорского семейства Венская императорская академия тщательно изучила ее наследование и опубликовала результаты, вместе с историческими портретами. Судя по всему, этот признак является истинно менделевской «аллелью» по отношению к нормальной форме губы. Сосредоточившись на портретах члена семейства, жившего в XVI веке, и его потомка, жившего в XIX веке, можно смело предположить, что материальная структура гена, ответственная за эту аномальную черту, передавалась из поколения в поколение, воспроизводимая при каждом из не слишком многочисленных клеточных делений, разделяющих этих двух людей. Более того, число атомов, вовлеченных в соответствующую генную структуру, скорее всего, имеет тот же порядок, что и в случаях, изученных с помощью рентгеновского излучения. Все это время ген находился при температуре около 98 °F[25]. Как объяснить, что за столетия разупорядочивающее тепловое движение не нарушило его целостности?
В конце прошлого века физик не смог бы ответить на данный вопрос, если бы опирался только на законы природы, которые был способен истолковать и действительно понимал. Поразмыслив о статистической ситуации, он бы сказал (как мы заметим, совершенно верно): этими материальными структурами могут быть только молекулы. К тому времени химия уже обладала обширными знаниями о существовании, а иногда и очень высокой стабильности таких атомных ассоциаций. Однако эти знания были чисто эмпирическими. Природа молекул оставалась загадкой – крепкая связь между атомами, поддерживающая форму молекулы, ставила всех в тупик. Да, это был бы верный ответ, однако его ценность была бы ограничена до тех пор, пока загадочная биологическая стабильность сводилась бы лишь к не менее загадочной химической стабильности. Свидетельство того, что два признака со схожим проявлением основаны на одном принципе, всегда шатко, если этот принцип неизвестен.
В данном случае объяснение дает квантовая теория. Согласно современным знаниям, механизм наследственности тесно связан – нет, основывается на самом базисе квантовой теории. Теорию эту открыл Макс Планк[26] в 1900 году. Современную генетику можно датировать подтверждением де Фризом, Корренсом и Чермаком статьи Менделя (1900) и статьей де Фриза о мутациях (1903). Таким образом, две великие теории возникли почти одновременно, и неудивительно, что обеим следовало достичь определенной зрелости, прежде чем между ними проявится связь. Квантовой теории на это потребовалось более четверти века, пока в 1926 году В. Гайтлер[27] и Ф. Лондон[28] не разработали общие принципы квантовой теории химической связи. Теория Гайтлера – Лондона включает тонкие и сложные концепции последних достижений квантовой теории (квантовую механику, или волновую механику). Представить ее без вычислений практически невозможно или потребует отдельной небольшой книги. Но, к счастью, теперь, когда вся работа по прояснению нашего мышления проделана, можно прямо обозначить связь между квантовыми переходами и мутациями, выделить наиболее примечательную черту. Этим мы и займемся.
Величайшим откровением квантовой теории стало открытие дискретности в природе, в контексте, где, согласно сложившимся взглядам, что-либо кроме непрерывности выглядит абсурдным.
Первый подобный случай касался энергии. Крупное тело меняет энергию непрерывно. Например, раскачанный маятник постепенно замедляется благодаря сопротивлению воздуха. Странно, но следует признать, что система атомных масштабов ведет себя иначе. По причинам, в которые мы не можем здесь углубиться, нужно полагать, что маленькая система по природе своей обладает определенными дискретными величинами энергии, их называют свойственными ей энергетическими уровнями. Переход из одного состояния в другое представляет собой весьма загадочное событие, обычно называемое квантовым переходом.
Однако энергия – не единственная характеристика системы. Снова рассмотрим наш маятник, но такой, что осуществляет различные движения, – тяжелый шар, подвешенный на нитке к потолку. Он может качаться в направлении север – юг, или восток – запад, или любом ином, или описывать круг, или эллипс. Аккуратно обдувая шар мехами, можно заставить маятник непрерывно переключаться от одного вида движения к другому.
Для мелкомасштабных систем большинство этих или схожих характеристик – мы не станем вдаваться в детали – меняются дискретно. Они «квантованы», как энергия.
В результате атомные ядра с их электронными оболочками, оказываясь в непосредственной близости друг к другу и формируя систему, не могут принять любую произвольную конфигурацию. По своей природе они выбирают из многочисленного, но дискретного числа состояний[29]. Обычно мы называем их уровнями, или энергетическими уровнями, поскольку энергия играет важную роль в данной характеристике. Но следует понимать, что полное описание включает не только энергию. По сути надо говорить о состоянии как об определенной конфигурации всех частиц.
Переход из одной конфигурации в другую является квантовым переходом. Если вторая конфигурация обладает большей энергией («более высокий уровень»), для перехода необходимо снабдить систему извне количеством энергии, хотя бы равным разнице между двумя уровнями. Переход на низший уровень может происходить самопроизвольно, а избыток энергии будет рассеян в виде излучения.
Среди дискретных состояний некой выборки атомов может существовать самый нижний уровень, соответствующий близкому расположению ядер относительно друг друга. В таком состоянии атомы формируют молекулу. Следует подчеркнуть, что молекула поневоле будет обладать определенной стабильностью. Ее конфигурация остается неизменной, если извне нет притока энергии, равной хотя бы разнице, необходимой для «подъема» на более высокий уровень. Таким образом, эта разница между уровнями, имеющая количественное выражение, количественно определяет степень стабильности молекулы. Мы увидим, как тесно данный факт связан с самими основами квантовой теории – с дискретностью энергетических уровней.
Я вынужден просить читателя принять на веру то, что этот набор идей полностью подтверждается химическими фактами – и успешно объясняет основополагающую концепцию химической валентности и многие детали молекулярной структуры, энергию связи, стабильность при различных температурах и т. п. Я говорю о теории Гайтлера – Лондона, которую, как я уже упоминал, невозможно рассмотреть здесь подробно.
Мы должны удовлетвориться рассмотрением вопроса, представляющего важность для нашего биологического исследования, а именно стабильности молекулы при различных температурах. Предположим, что наша система атомов вначале находится в состоянии с минимальной энергией. Физик назовет это молекулой при абсолютном нуле температуры. Чтобы перевести ее на следующий более высокий уровень, необходим приток определенного количества энергии. Простейший способ предоставить его – «нагреть» молекулу, перенести ее в окружающую среду с большей температурой (термостат), тем самым позволив другим системам (атомам, молекулам) сталкиваться с нашей молекулой. Поскольку тепловое движение хаотично, не существует температурного порога, при котором «подъем» произойдет обязательно и немедленно. Скорее при любой температуре, отличной от абсолютного нуля, существует бо́льшая или меньшая вероятность такого «подъема», и она, разумеется, растет с ростом температуры в термостате. Лучший способ выразить эту вероятность – указать среднее время, которое придется ждать перехода, – «время ожидания».