Человек: общие данные
В процессе изучения человека его структуры условно подразделяют на клетки, ткани, органы, системы и аппараты органов, которые и формируют организм. Однако следует предостеречь читателя от буквального понимания этого деления. Организм един, он может существовать лишь благодаря своей целостности, но в нем выделяют ряд иерархических уровней: клеточный, тканевый, органный, системный, организменный (табл. 1).
Клетка
Клетка является основной структурной и функциональной единицей живых организмов, осуществляющей рост, развитие, обмен веществ и энергии, хранящейся, перерабатывающей и реализующей генетическую информацию. Клетка представляет собой сложную систему биополимеров, отделенную от внешней среды плазматической мембраной (цитолеммой, плазмалеммой) и состоящую из ядра и цитоплазмы, в которой располагаются органеллы и включения.
Немецкий ученый Т. Шванн в середине XIX века создал клеточную теорию, основные положения которой свидетельствовали о том, что все ткани и органы состоят из клеток; клетки растений и животных принципиально сходны между собой, все они возникают одинаково; деятельность организмов – сумма жизнедеятельности отдельных клеток. Большое влияние на дальнейшее развитие клеточной теории и вообще на учение о клетке оказал великий немецкий ученый Р. Вирхов. Он не только свел воедино все многочисленные разрозненные факты, но и убедительно показал, что клетки являются постоянной структурой и возникают только путем размножения.
Клеточная теория в современной интерпретации включает в себя следующие главные положения: клетка является универсальной элементарной единицей живого; клетки всех организмов принципиально сходны по своему строению, функции и химическому составу; клетки размножаются только путем деления исходной клетки; многоклеточные организмы являются сложными клеточными ансамблями, образующими целостные системы. Благодаря современным методам исследования были выявлены два основных типа клеток: более сложно организованные, высокодифференцированные эукариотические клетки (растения, животные и некоторые простейшие, водоросли, грибы и лишайники) и менее сложно организованные прокариотические клетки (сине-зеленые водоросли, актиномицеты, бактерии, спирохеты, микоплазмы, риккетсии, хламидии). В отличие от прокариотической эукариотическая клетка имеет ядро, ограниченное двойной ядерной мембраной, и большое количество мембранных органелл.
Таблица 1
Иерархические уровни строения организма

ВНИМАНИЕ
Клетка является основной структурной и функциональной единицей живых организмов, осуществляющей рост, развитие, обмен веществ и энергии, хранящей, перерабатывающей и реализующей генетическую информацию.
С точки зрения морфологии клетка представляет собой сложную систему биополимеров, отделенную от внешней среды плазматической мембраной (плазмолеммой) и состоящую из ядра и цитоплазмы, в которой располагаются органеллы и включения (гранулы) (рис. 1). Клетки разнообразны по своей форме, строению, химическому составу и характеру обмена веществ. Все клетки гомологичны, т. е. имеют ряд общих структурных признаков, от которых зависит выполнение основных функций. Клеткам присуще единство строения, метаболизма (обмена веществ) и химического состава. Вместе с тем различные клетки имеют и специфические структуры. Это связано с выполнением ими специальных функций.
Химический состав клетки
В состав клетки входит более 100 химических элементов, на долю четырех из них приходится около 98 % массы, это органогены: кислород (65–75 %), углерод (15–18 %), водород (8–10 %) и азот (1,5–3,0 %). Остальные элементы подразделяются на три группы: макроэлементы – их содержание в организме превышает 0,01 %); микроэлементы (0,00001–0,01 %) и ультрамикроэлементы (менее 0,00001). К макроэлементам относятся сера, фосфор, хлор, калий, натрий, магний, кальций. К микроэлементам – железо, цинк, медь, йод, фтор, алюминий, медь, марганец, кобальт и др. К ультрамикроэлементам – селен, ванадий, кремний, никель, литий, серебро и до. Несмотря на очень малое содержание, микроэлементы и ультрамикроэлементы играют очень важную роль. Они влияют, главным образом, на обмен веществ. Без них невозможна нормальная жизнедеятельность каждой клетки и организма как целого.
Клетка состоит из неорганических и органических веществ. Среди неорганических наибольшее количество воды. Относительное количество воды в клетке составляет от 70 до 80 %. Вода – универсальный растворитель, в ней происходит все биохимические реакции в клетке. При участии воды осуществляется теплорегуляция. Вещества, растворяющиеся в воде (соли, основания, кислоты, белки, углеводы, спирты и др.), называются гидрофильными. Гидрофобные вещества (жиры и жироподобные) не растворяются в воде. Другие неорганические вещества (соли, кислоты, основания, положительные и отрицательные ионы) составляют от 1,0 до 1,5 %.

Рис. 1. Ультрамикроскопическое строение клетки
1 – цитолемма (плазматическая мембрана); 2 – пиноцитозные пузырьки; 3 – центросома клеточный центр (цитоцентр); 4 – гиалоплазма; 5 – эндоплазматическая сеть: а – мембрана зернистой сети; б – рибосомы; 6 – связь перинуклеарного пространства с полостями эндоплазматической сети; 7 – ядро; 8 – ядерные поры; 9 – незернистая (гладкая) эндоплазматическая сеть; 10 – ядрышко; 11 – внутренний сетчатый аппарат (комплекс Гольджи); 12 – секреторные вакуоли; 13 – митохондрия; 14 – липосомы; 15 – три последовательные стадии фагоцитоза; 16 – связь клеточной оболочки (цитолеммы) с мембранами эндоплазматической сети
Среди органических веществ преобладают белки (10–20 %), жиры, или липиды (1–5 %), углеводы (0,2–2,0 %), нуклеиновые кислоты (1–2 %). Содержание низкомолекулярных веществ не превышает 0,5 %.
Молекула белка является полимером, который состоит из большого количества повторяющихся единиц мономеров. Мономеры белка аминокислоты (их 20) соединены между собой пептидными связями, образуя полипептидную цепь (первичную структуру белка). Она закручивается в спираль, образуя, в свою очередь, вторичную структуру белка. Благодаря определенной пространственной ориентации полипептидной цепи возникает третичная структура белка, которая определяет специфичность и биологическую активность молекулы белка. Несколько третичных структур, объединяясь между собой, образуют четвертичную структуру.
Белки, входя в состав всех клеточных структур, выполняют пластическую (строительную) функцию. Движения клеток также осуществляют белки. Они обеспечивают транспорт веществ в клетку, из клетки и внутри клетки. Важной является защитная функция белков (антитела). Белки являются одним из источников энергии. Ферменты являются белками.
Углеводы подразделяются на моносахариды и полисахариды. Последние построены из моносахаридов, являющихся, подобно аминокислотам, мономерами. Среди моносахаридов в клетке наиболее важны глюкоза, фруктоза (содержит шесть атомов углерода) и пентоза (пять атомов углерода). Пентозы входят в состав нуклеиновых кислот. Моносахариды хорошо растворяются в воде. Полисахариды плохо растворяются в воде (в животных клетках гликоген, в растительных – крахмал и целлюлоза. Углеводы являются источником энергии, сложные углеводы, соединенные с белками (гликопротеиды), жирами (гликолипиды), участвуют в образовании клеточных поверхностей и взаимодействиях клеток.
К липидам относятся жиры и жироподобные вещества. Молекулы жиров построены из глицерина и жирных кислот. К жироподобным веществам относятся холестерин, некоторые гормоны, лецитин. Липиды, являющиеся основным компонентом клеточных мембран (они описаны ниже), выполняют тем самым строительную функцию. Липиды – важнейшие источники энергии. Так, если при полном окислении 1 г белка или углеводов освобождается 17,6 кДж энергии, то при полном окислении 1 г жира – 38,9 кДж. Липиды осуществляют терморегуляцию, защищают органы (жировые капсулы).
Нуклеиновые кислоты являются полимерными молекулами, образованными мономерами нуклеотидами. Нуклеотид состоит из пуринового или пиримидинового основания, сахара (пентозы) и остатка фосфорной кислоты. Во всех клетках существует два типа нуклеиновых кислот: дезоксирибонулеиновая (ДНК) и рибонуклеиновая (РНК), которые отличаются по составу оснований и сахаров (табл. 2, рис. 2).
Молекула ДНК состоит из двух полинуклеотидных цепей, закрученных одна вокруг другой в виде двойной спирали. Азотистые основания обеих цепей соединены между собой комплементарно водородными связями. Аденин соединяется только с тимином, а цитозин – с гуанином (А – Т, Г – Ц). В ДНК записана генетическая информация, которая определяет специфичность синтезируемых клеткой белков, т. е. последовательность аминокислот в полипептидной цепи. ДНК передает по наследству все свойства клетки. ДНК содержится в ядре и митохондриях.

Рис. 2. Пространственная структура нуклеиновых кислот (по Б. Албертсу и соавт., с изм.)
I – РНК; II – ДНК; ленты – сахарофосфатные остовы; A, C, G, T, U – азотистые основания, решетки между ними – водородные связи
Молекула РНК образована одной полинуклеотидной цепью. В клетках существует три типа РНК. Информационная, или мессенджер РНК тРНК (от англ. messenger – «посредник»), которая переносит информацию о нуклеотидной последовательности ДНК в рибосомы (см. ниже).
Транспортная РНК (тРНК), которая переносит аминокислоты в рибосомы. Рибосомальная РНК (рРНК), которая участвует в образовании рибосом. РНК содержится в ядре, рибосомах, цитоплазме, митохондриях, хлоропластах.
Таблица 2
Состав нуклеиновых кислот

Строение клетки человека
Для всех клеток типично наличие цитоплазмы и ядра (см. рис. 1). Цитоплазма включает в себя гиалоплазму. В клетках встречаются также временные клеточные структуры включения.
Размеры клеток человека варьируют от нескольких микрометров[1] (например, малый лимфоцит) до 200 мкм (яйцеклетка). В организме человека встречаются клетки различной формы: овоидные, шаровидные, веретеновидные, плоские, кубические, призматические, полигональные, пирамидальные, звездчатые, чешуйчатые, отросчатые, амебовидные.
Снаружи каждая клетка покрыта плазматической мембраной (плазмалеммой) толщиной 9–10 нм, ограничивающей клетку от внеклеточной среды. Они выполняет следующие функции: транспортную, защитную, разграничительную, рецепторную восприятия сигналов внешней (для клетки) среды, участие в иммунных процессах, обеспечение поверхностных свойств клетки.
Будучи очень тонкой, плазмалемма не видна в световом микроскопе. В электронном микроскопе, если срез проходит под прямым углом к плоскости мембраны, последняя представляет собой трехслойную структуру, наружная поверхность которой покрыта тонкофибриллярным гликокаликсом толщиной от 75 до 2000 А°, совокупность молекул, связанных с белками плазмолеммы.
Таблица 3
Структурные компоненты клетки

Плазмалемма, как и другие мембранные структуры, состоит из двух слоев амфипатических[2] молекул липидов (билипидный слой, или бислой). Их гидрофильные «головки» направлены к наружной и внутренней сторонам мембраны, а гидрофобные «хвосты» обращены друг к другу В билипидный слой погружены молекулы белка. Некоторые из них (интегральные, или внутренние трансмембранные белки) проходят через всю толщу мембраны, другие (периферические, или внешние) лежат во внутреннем или наружном монослое мембраны. Некоторые интегральные белки связаны нековалентными связями с белками цитоплазмы (рис. 3). Подобно липидам, белковые молекулы также являются амфипатическими их гидрофобные участки окружены аналогичными «хвостами» липидов, а гидрофильные обращены наружу или внутрь клетки или в одну сторону.
ВНИМАНИЕ
Белки осуществляют большую часть мембранных функций: многие мембранные белки являются рецепторами, другие ферментами, третьи переносчиками.
Плазмалемма образует ряд специфических структур. Это межклеточные соединения, микроворсинки, реснички, клеточные инвагинации и отростки.
Микроворсинки – это лишенные органелл пальцевидные выросты клетки, покрытые плазмалеммой, длиной 1–2 мкм и диаметром до 0,1 мкм. Некоторые эпителиальные клетки (например, кишечные) имеют очень большое количество микроворсинок, образуя так называемую щеточную каемку. Наряду с обычными микроворсинками на поверхности некоторых клеток имеются крупные микроворсинки стереоцилии (например, волосковые сенсорные клетки органов слуха и равновесия эпителиоциты протока придатка яичка и др.).

Рис. 3. Строение клеточной мембраны, схема (по А. Хэму и Д. Кормаку)
1 – углеводные цепи; 2 – гликолипид; 3 – гликопротеид; 4 – углеводородный «хвост»; 5 – полярная «головка»; 6 – белок; 7 – холестерин; 8 – микроктрубочки
Немембранные органеллы
Реснички и жгутики выполняют функцию движения. До 250 ресничек длиной 5–15 мкм диаметром 0,15–0,25 мкм покрывают апикальную поверхность эпителиальных клеток верхних дыхательных путей, маточных труб, семявыводящих канальцев. Ресничка представляет собой вырост клетки, окруженный плазмолеммой. В центре реснички проходит осевая нить, или аксонема, образованная 9 периферическими дуплетами микротрубочек, окружающих одну центральную пару. Периферические дуплеты, состоящие из двух микротрубочек, окружают центральную капсулу. Периферические дуплеты заканчиваются в базальном тельце (кинетосоме), которое образовано из 9 триплетами микротрубочек. На уровне плазмалеммы апикальной части клетки триплеты переходят в дуплеты, здесь же начинается и центральная пара микротрубочек (рис. 4). Жгутики эукариотических клеток напоминают реснички. Реснички совершают координированные колебательные движения.

Рис. 4. Ресничка
I – ресничка на продольно-поперечном разрезе; II – внеклеточная часть; III – поперечный срез через внеклеточную часть; IV – поперечный срез через промежуточную область, соединяющую аксонему с базальным тельцем; V – срез через базальное тельце; 1 – ресничка; 2 – периферические дуплеты; 3 – центральный дуплет; 4 – центральная капсула; 5 – микроворсинки; 6 – наружные микротрубочки; 7 – базальное тельце; 8 – базальный корешок; 9 – цитолемма; 10 – периферические дуплеты; 11 – центральный дуплет; 12 – центральная капсула; 13 – спица; 14 – субфибрилла а; 15 – субфибрилла б; 16 – динеин; 17 – триплеты микротрубочек; 18 – триплеты;19 – спицы; 20 – центральный цилиндр
Клеточный центр, образованный двумя центриолями (диплосома), находится вблизи ядра, расположенными под углом друг к другу (рис. 5). Каждая центриоль представляет собой цилиндр, стенка которого состоит из 9 триплетов микротрубочек длиной около 0,5 мкм и диаметром около 0,25 мкм. Триплеты, расположенные по отношению друг к другу под углом около 50°, состоят из трех микротрубочек. Центриоли удваиваются в клеточном цикле. Не исключено, что, подобно митохондриям, центриоли содержат собственную ДНК. Центриоли участвуют в образовании базальных телец ресничек и жгутиков и в образовании митотического веретена.

Рис. 5. Клеточный центр и другие структуры цитоплазмы, схема (по Р. Крстичу, с изм.)
1 – центросфера; 2 – центриоль на поперечном срезе (триплеты микротрубочек, радиальные спицы, центральная структура «колеса телеги»); 3 – центриоль (продольный разрез); 4 – сателлиты; 5 – окаймленные пузырьки; 6 – зернистая эндоплазматическая сеть; 7 – митохондрия; 8 – внутренний сетчатый аппарат (комплекс Гольджи); 9 – микротрубочки
Цитоскелет клетки представляет собой трехмерную сеть, в которой различные органеллы и растворимые белки связаны с микротрубочками. Главную роль в образовании цитоскелета играют микротрубочки, помимо них принимают участие актиновые, миозиновые и промежуточные филаменты. Микротрубочки, имеющиеся в цитоплазме всех эукариотических клеток, образованы белком тубулином. Микротрубочки образуют клеточный скелет (цитоскелет) и участвуют в транспорте веществ внутри клетки (рис. 6).

Рис. 6. Микротрубочка
1 – тубулиновые субъединицы; 2 – ассоциированные белки; 3 – перемещаемые частицы
Мембранные органеллы. Транспорт через мембраны
Для клеток человека характерно наличие огромного количества внутриклеточных мембран, образующих несколько компартментов (от англ. compartment – «отделение, купе»), отличающихся друг от друга строением и функцией: цитозоль, ядро, эндоплазматический ретикулум, комплекс Гольджи, митохондрии, лизосомы, пероксисомы. Благодаря наличию этих элементов в клетке одновременно протекает большое количество различных биохимических реакций.
Все мембранные органеллы построены из элементарных мембран, принцип строения которых аналогичен описанному выше строению плазмалеммы. Поглощение клетками макромолекул и частиц происходит путем эндоцитоза (от греч. endon – «внутри», kytos – «клетка»), выделение – путем экзоцитоза (от греч. ехо – «вне», kytos – «клетка»).
Одной из важнейших функций плазмолеммы является транспорт. Напомним, что обращенные друг к другу гидрофобные «хвосты» липидов препятствуют проникновению полярных водорастворимых молекул. Различают два вида транспорта: пассивный и активный. Первый не требует затрат энергии, второй энергозависимый. Как правило, внутренняя (цитоплазматическая) поверхность мембраны несет отрицательный заряд, что облегчает проникновение в клетку положительно заряженных ионов. Вода поступает в клетку путем осмоса (от греч. osmos – «толчок, давление»), который представляет собой медленное проникновение воды через полупроницаемую мембрану, разделяющую два раствора различной концентрации. В результате концентрация этих двух растворов выравнивается.
Диффузия (от лат. diffusion – «распространение, растекание») – это переход ионов или молекул, вызванный их броуновским движением через мембраны из зоны, где эти вещества находятся в более высокой концентрации, в зону с более низкой концентрацией до тех пор, пока концентрации по обе стороны мембраны выравниваются. Специфические транспортные белки, встроенные в мембрану, переносят через нее небольшие полярные молекулы, причем каждый белок осуществляет транспорт одного класса молекул или только одного соединения. Некоторые трансмембранные белки образуют каналы. Активный транспорт осуществляют белки-переносчики, при этом расходуется энергия, получаемая вследствие гидролиза АТР (аденозинтрифосфатной кислоты) или протонного потенциала. Активный транспорт происходит против градиента концентрации. Для осуществления биохимических реакций необходимо поступление веществ в клетку путем эндоцитоза и выведение продуктов обмена экзоцитоза.
Эндоцитоз. Известны две разновидности эндоцитоза: фагоцитоз – поглощение твердых частиц (от греч. phagos – «пожирающий» и kytos – «клетка») и пиноцитоз – поглощение растворенных веществ (от греч. Pino – «пью»). Фагоцитированная частичка, заключенная в мембрану, называется фагосомой. В процессе эндо- и экзоцитоза транспортируемые вещества заключены в мембранные пузырьки. Для того чтобы внешние молекулы поступили в клетку, они должны быть сначала связаны рецепторами гликокаликса. Цитолемма начинает впячиваться, затем ее края сближаются и смыкаются, отщепляя пузырек, не сущий в себе захваченные молекулы. Образуется эндосома, которая погружается в цитоплазму и встречается с лизосомами. Их мем браны сливаются. В возникшей вторичной лизосоме вещества, по ступившие в клетку, подвергаются расщеплению.
Экзоцитоз обеспечивает выведение крупномолекулярных соединений. Сначала они обособляются в комплексе Гольджи в виде транспортных пузырьков и направляются к клеточной поверхности. Мембрана пузырька встраивается в плазмалемму, и содержимое пузырька оказывается за пределами клетки.
Эндоплазматическая сеть, или эндоплазматический ретикулум (ЭР), представляет собой единую непрерывную полость, ограниченную мембраной, образующей множество инвагинаций и складок (рис. 7). Поэтому на электронограммах эндоплазматическая сеть выглядит в виде множества трубочек, плоских или округлых цистерн, мембранных пузырьков. Различают два типа ЭР: гранулярный и агранулярный. Обращенная к цитозолю сторона первого покрыта рибосомами, второго лишена их. Функция гранулярного ЭР: синтез белков рибосомами и транспорт белков, гладкого синтез и обмен углеводов и липидов (стероидных гормонов, гликогена, холестерола) и обезвреживание (гепатоциты), синтез хлоридов, из которых в желудке образуется соляная кислота. Будучи депо ионов кальция, гладкий ЭР участвует в мышечном сокращении; отграничивает будущие тромбоциты в мегакариоцитах. Одной из важнейших функций ЭР является синтез мембранных белков и липидов для всех клеточных органелл.

Рис. 7. Эндоплазматическая сеть (по Крстичу, с изм.)
1 – трубочки гладкой (агранулярной) эндоплазматической сети; 2 – цистерны шероховатой гранулярной сети; 3 – поровый комплекс; 4 – наружная ядерная мембрана, покрытая рибосомами; 5 – внутренняя ядерная мембрана
Комплекс, или аппарат Гольджи (КГ), представляет собой совокупность цистерн, пузырьков, пластинок, трубочек, мешочков, ограниченных мембраной, в которых накапливаются и упаковываются синтезированные продукты (рис. 8). Эти продукты с помощью элементов комплекса выводятся из клетки, кроме того, в них происходит синтез полисахаридов, образование белково-углеводных комплексов и модификация переносимых молекул. В световом микроскопе КГ выглядит в виде сеточки или системы канальцев и вакуолей. КГ имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток КГ расположен вокруг или вблизи ядра. В КГ выявляются три мембранных элемента: уплощенные мешочки (цистерны), пузырьки и вакуоли. КГ – трехмерная структура чашеобразной формы, состоящая из нескольких (от одной до нескольких сот) диктиосом (от греч. dyktion – «сеть»). Каждая диктиосома содержит 4–8 (в среднем 6) лежащих параллельно уплощенных цистерн, пронизанных порами с расширенными концами, от которых отщепляются вакуоли, содержащие синтезированные вещества. Цистерны ассоциированы со множеством мембранных пузырьков, а также с более крупными секреторными гранулами. Элементы комплекса Гольджи связаны между собой каналами.
Мембраны комплекса Гольджи образуются и поддерживаются гранулярным эндоплазматическим ретикулумом, в котором синтезируются мембранные компоненты. Они переносятся транспортными пузырьками, отпочковывающимися от ЭР, и сливаются с КГ, от которого постоянно отпочковываются секреторные пузырьки, а мембраны цистерн постоянно обновляются. Они поставляют гликокаликс и синтезированные вещества к плазмолемме, таким образом обеспечивается и обновление плазмолеммы. Одной из важнейших функций КГ является сортировка белков.

Рис. 8. Комплекс Гольджи (по Крстичу, с изм.)
1 – транспортные пузырьки; 2 – цистерна гранулярной ЭПС; 3 – транспортные пузырьки, отпочковывающиеся от гранулярной ЭПС; 4 – слияние транспортного пузырька с формирующейся поверхностью комплекса Гольджи; 5 – конденсирующие вакуоли; 6 – пузырьки, отщепляющиеся от комплекса Гольджи; 7 – формирующаяся поверхность комплекса Гольджи; 8 – зрелая поверхность комплекса Гольджи
Лизосомы – мембранные органеллы, содержащие около 50 видов различных гидролитических ферментов, которые синтезируются на рибосомах гранулярного эндоплазматического ретикулума, откуда переносятся транспортными пузырьками в КГ, где они видоизменяются. От поверхности КГ отпочковываются первичные лизосомы. Все лизосомы клетки формируют единое лизосомное пространство, в котором постоянно поддерживается кислая среда рН колеблется в пределах 3,5–5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия.
Различают четыре функциональные формы лизосом (рис. 9.). Первичные лизосомы, отпочковавшиеся от комплекса Гольджи, сливаясь с фагосомой, образуют вторичную лизосому (фаголизосому), в которой происходит переваривание поглощенных веществ до мономеров. Последние транспортируются через лизосомальную мембрану в цитозоль. Непереваренные вещества остаются в лизосоме, в результате чего образуется остаточное тельце. Кроме того, лизосомы переваривают поврежденные структуры собственной клетки (аутолизосома).

Рис. 9. Строение и функционирование лизосом (возможные пути формирования вторичных лизосом путем слияния мишеней с первичными лизосомами, содержащими новосинтезированные гидролитические ферменты); стрелки – направления движения (по де Дюву и по Албертсу и соавт., с изм.)
1 – рециклирование мембран; 2 – цитолемма; 3 – кринофагия; 4 – фагоцитоз; 5 – вторичная лизосома; 6 – фагосома; 7 – остаточное тельце; 8 – мультивезикулярное тельце; 9 – очистка лизосом от мономеров; 10 – пиноцитоз; 11 – аутофагосома; 12 – начало аутофагии; 13 – участок агранулярной эндоплазматической сети; 14 – гранулярная эндоплазматическая сеть; 15 – протонный насос; 16 – первичные лизосомы; 17 – комплекс Гольджи
Пероксисомы представляют собой пузырьки диаметром от 0,2 до 0,5 мкм, окруженные мембранами, содержащие окислительные ферменты (около 40 % всех белков составляет каталаза), производящие и разрушающие перекись водорода. Они используют молекулярный кислород.
Митохондрии, являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в форму, доступную для использования клеткой. В световом микроскопе митохондрии выглядят в виде округлых, удлиненных или палочковидных структур длиной 0,3–5,0 мкм и шириной 0,2–1,0 мкм. Количество, размеры и расположение митохондрий зависят от функции клетки, ее потребности в энергии. Так, в каждой печеночной клетке их количество достигает 2500. С помощью электронной микроскопии установлено, что митохондрии являются органеллами с двойными мембранами (рис. 10). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство. Внутренняя мембрана образует многочисленные складки, или кристы, благодаря которым внутренней мембраны резко возрастает. На внутренней поверхности крист лежит множество электронноплотных субмитохондриальных элементарных частиц (до 4000 на 1 мкм2 мембраны), имеющих форму гриба. В пространстве, ограниченном внутренней митохондриальной мембраной, находится мелкозернистый матрикс.
Митохондрии содержат собственную ДНК, РНК и рибосомы, которые находятся в матриксе. Таким образом, митохондрии снабжены собственной генетической системой, необходимой для их самовоспроизведения и синтеза белков. Следует подчеркнуть, что митохондриальные ДНК, РНК и рибосомы отличаются от таковых собственной клетки и весьма сходны с прокариотическими.

Рис. 10. Митохондрия (по Б. Албертсу и др.; по К. де Дюву, с изм.)
I – общая схема строения: 1 – наружная мембрана; 2 – внутренняя мембрана; 3 – кристы; 4 – матрикс; II – схема строения кристы: 5 – складка внутренней мембраны; 6 – грибовидные тельца
ВНИМАНИЕ
У млекопитающих, в том числе и человека, митохондриальный геном наследуется от матери.
Митохондрии размножаются путем деления уже существующих независимо от деления других митохондрий и самой клетки.
В клетках постоянно происходит метаболизм (от греч. metabole – «перемена, превращение»), или обмен веществ, который представляет собой совокупность процессов ассимиляции (реакции биосинтеза сложных биологических молекул из более простых) и диссимиляции (реакции расщепления). В результате диссимиляции освобождается энергия, заключенная в химических связях веществ. Эта энергия используется клеткой для осуществления различной работы, в том числе и ассимиляции. Напомним, что энергия не возникает и не уничтожается, она лишь переходит из одного вида в другой, пригодный для выполнения работы. Клетка использует энергию, заключенную в химических связях аминокислот, моносахаридов и жирных кислот. Они образуются в результате пищеварения из белков, углеводов и жиров и поступают в клетку.
Рассмотрим энергетический обмен на примере расщепления глюкозы. Глюкоза транспортируется через плазматическую мембрану, и в цитоплазме происходит ее бескислородное расщепление, или гликолиз. Гликолиз – это многоступенчатый ферментативный процесс, в результате которого из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты и две молекулы АТР (с учетом двух молекул АТР, затрачиваемых для осуществления реакций). Пировиноградная кислота подвергается дальнейшему окислению (аэробному при участии кислорода) в митохондриях, в которых имеются цепи ферментов, катализирующие реакции синтеза АТР (аденозинтрифосфат). АТР является универсальным переносчиком и основным аккумулятором энергии в клетке. Энергия заключена в высокоэнергетических связях между остатками фосфорной кислоты.
При отщеплении от АТР одной фосфатной группы образуются АДР (аденозиндифосфатная кислота) и фосфат и выделяется свободная энергия, которая используется клеткой для осуществления работы. В митохондриях АДР, соединяясь с остатком фосфорной кислоты, превращается в АТР. В результате гликолиза освобождается лишь около 5 % энергии, остальная освобождается в митохондриях в процессе аэробного окисления и запасается в АТР. В расчете на одну молекулу глюкозы образуется 36 молекул АТР.
Ядро – основная клеточная структура, имеется во всех клетках человека, кроме эритроцитов и тромбоцитов. В большинстве клеток его форма шаровидная или овоидная, однако встречаются и другие формы ядра (кольцевидное, палочковидное, веретеновидное, четковидное, бобовидное, сегментированное, полиморфное и др.). Размеры ядер колеблются в широких пределах от 3 до 25 мкм. Наиболее крупное ядро имеет яйцеклетка. Большинство клеток человека одноядерные, однако имеются двухъядерные (например, некоторые нейроны, гепатоциты, кардиомиоциты), а некоторые структуры многоядерные (мышечные волокна миосимпласты).
В ядре различают следующие структуры: ядерную оболочку, хроматин, ядрышко и нуклеоплазму. Ядро окружено ядерной оболочкой, состоящей из внутренней и наружной ядерных мембран толщиной 8 нм каждая, разделенных перинуклеарным пространством (или цистерной ядерной оболочки) шириной 20–50 нм. Обе являются элементарными клеточными мембранами. К наружной, переходящей в гранулярный эндоплазматический ретикулум, прикреплены рибосомы. Перинуклеарное пространство составляет единую полость с эндоплазматическим ретикулумом (рис. 11).
Ядерная оболочка пронизана множеством расположенных упорядоченно ядерных пор округлой формы диаметром 50–70 нм, которые в общей сложности занимают до 25 % поверхности ядра. Через ядерные поры осуществляется избирательный транспорт крупных частиц, а также обмен веществ между ядром и цитозолем.

Рис. 11. Ядро (по Албертсу и др., с изм.)
1 – наружная мембрана кариотеки (наружная ядерная мембрана); 2 – перинуклеарное пространство; 3 – внутренняя мембрана кариотеки (внутренняя ядерная мембрана); 4 – ядерная пластинка; 5 – поровый комплекс; 6 – рибосомы; 7 – нуклеоплазма; 8 – гетерохроматин; 9 – цистерна зернистой эндоплазматической сети; 10 – эухроматин; 11 – ядрышко
В живых клетках кариоплазма (нуклеоплазма) гомогенна (кроме ядрышка). После фиксации и обработки тканей для световой или электронной микроскопии в ней становятся видными два типа хроматина (от греч. chroma – «краска»); хорошо окрашивающийся гетерохроматин (неактивный) и светлый эухроматин (активный).
Каждая хромосома образована одной длинной молекулой дезоксинуклеопротеида, который представляет собой двойную цепь ДНК, окруженную сложной системой белков. В результате многократной упаковки образуются удлиненные палочковидные структуры, имеющие два плеча, разделенные центромерой (рис. 12).

Рис. 12. Уровни упаковки ДНК в хромосоме (по Албертсу и др., с изм.)
I – двойная спираль молекулы ДНК; II – нуклеосомная нить; 1 – гистон Н1; 2 – ДНК; 3 – прочие гистоны; III – хроматиновая фибрилла; IV – серия петельных доменов; V – конденсированный хроматин в составе петельного домена; VI – метафазная хромосома; 4 – центромера; 5 – сестринские хроматиды
В делящемся ядре хроматин спирализуется, образуя хромосомы. Хроматин неделящегося ядра и хромосомы делящегося образованы дезоксирибонуклеиновой кислотой (ДНК), связанной с РНК и белками (гистонами и негистоновыми). Следует подчеркнуть химическое тождество хроматина и хромосом.
В соматических клетках имеются по две копии каждой хромосомы, их называют гомологичными. Они одинаковы по длине, форме, строению, расположению полос и несут одни и те же гены, которые локализованы одинаково. Нормальный кариотип человека (от греч. karyon – «ядро ореха», typos – «образец») включает 22 пары аутосом и одну пару половых хромосом (ХХ женщины или XV мужчины) (рис. 13).

Рис. 13. Кариотип человека (здорового мужчины)
I – кариотип, общий вид; II – метафазные хромосомы
Ген участок ДНК, характеризующийся определенной последовательностью нуклеотидов, ответственный за синтез специфического белка. Ген является элементарной единицей наследственности.
Ядрышко (одно или несколько) выявляется во всех неделящихся ядрах в виде плотного интенсивно окрашивающего округлого однородного базофильного тельца, величина которого пропорциональна интенсивности белкового синтеза. В ядрышке образуются рибосомы. Ядерный сок – не окрашивающаяся электронносветлая часть ядра – коллоидный раствор белков, окружающий хроматин и ядрышко.
Клеточный цикл
Центральная догма современной биологии характеризует жизнь следующим образом.
Иными словами, наследственная информация, заключенная в ДНК, передается по наследству благодаря репликации (удвоению) ДНК. Генетическая информация, записанная в виде последовательности нукдеотидов ДНК, в процессе транскрипции переписывается в нуклеотидную последовательность РНК, которая, в свою очередь, определяет последовательность аминокислот соответствующей белковой молекулы.
Клеточный цикл представляет собой совокупность процессов, происходящих в клетке при подготовке ее к делению и во время собственно деления. Клеточный цикл (рис. 14) подразделяется на митоз и интерфазу, которая представляет собой промежуток времени между окончанием одного митоза и началом следующего. В интерфазе совершается главное событие репликация ДНК. Репликация (от лат. replicatio – «повторение») – это процесс передачи хранящейся в родительской ДНК генетической информации дочерней путем точного ее воспроизведения. При этом каждая родительская цепь ДНК является матрицей для синтеза дочерней. Перед началом удвоения две цепи ДНК начинают раскручиваться и расходиться. Вдоль каждой цепи комплементарно строится новая цепь, при этом напротив тимина родительской цепи к синтезируемой новой цепи добавляется аденин, напротив гуанина – цитозин, и оба основания соединяются водородными связями. Процесс заканчивается образованием двух идентичных двухцепочечных молекул ДНК, обе они идентичны материнской. В результате полуконсервативной репликации каждая из двух дочерних молекул ДНК состоит из одной старой и одной новой цепи. В интерфазе наиболее интенсивно синтезируется также РНК и белки, связанные с ДНК, и удваиваются центриоли. К концу интерфазы хроматин конденсирован, ядрышко хорошо видно, ядерная оболочка не повреждена, органеллы не изменены.

Рис. 14. Строение и репликации ДНК, схема
Митоз (от греч. mitos – «нить») животных клеток впервые был описан В. Флеммингом в 1882 г.
ВНИМАНИЕ
Митоз обеспечивает генетическую стабильность, образование новых клеток, увеличение их числа в организме и, следовательно, рост организма, а также процессы регенерации.
Клетка вступает в митоз, имея удвоенное (в интерфазе) число хромосом (46 × 2), т. е. в ядре находятся 46 d-хромосом, каждая из которых состоит из двух хроматид (s-хромосом). Митоз подразделяется на профазу, метафазу анафазу и телофазу (рис. 15).
В профазе хромосомы становятся различимыми под микроскопом. В конце профазы обе пары центриолей начинают расходиться к полюсам клетки. Одновременно возникает двухполюсное митотическое веретено, состоящее из микротрубочек. В метафазе разрушается ядерная оболочка, хромосомы выстраиваются в ряд по экватору веретена, а их центромеры прикрепляются к микротрубочкам веретена. Метафазная хромосома состоит из двух соединенных центромерой сестринских хроматид, каждая из которых содержит одну молекулу ДНК, уложенную в виде суперспирали.
В анафазе сестринские хроматиды разделяются и становятся отдельными s-хромосомами, которые расходятся к полюсам с одинаковой скоростью (около 1 мкм/мин).
В телофазе разделившиеся группы хромосом подходят к полюсам, разрыхляются, деконденсируются, переходя в хроматин, становятся активными. Примерно в середине телофазы начинается образование ядрышка, к концу телофазы восстанавливается ядерная оболочка в каждой дочерней клетке. Еще в конце анафазы плазматическая мембрана как бы инвагинируется, образуя борозду деления, которая углубляется. Дочерние клетки расходятся (рис. 16).

Рис. 15. Стадии митоза. Показаны конденсация хроматина с образованием хромосом, образование веретена деления и равномерное распределение хромосом и центриолей по двум дочерним клеткам (по А. Хэму и Д. Кормаку, с изменениями)
А – интерфаза; Б – профаза; В – метафаза; Г – анафаза; Д – телофаза; Е – поздняя телофаза; 1 – ядрышко, 2 – центриоли; 3 – веретено деления; 4 – звезда; 5 – ядерная оболочка; 6 – кинетохор; 7 – непрерывные микротрубочки; 8, 9 – хромосомы; 10 – хромосомные микротрубочки; 11 – формирование ядра; 12 – борозда дробления; 13 – пучок актиновых нитей; 14 – остаточное (срединное) тельце
Мейоз. В жизненном цикле человека и других организмов, размножающихся половым путем, имеются два поколения постоянно чередующихся клеток: диплоидных (соматических) и гаплоидных (половых). Мейоз (от греч. meiosis – «уменьшение»), впервые открытый у животных В. Флеммингом в 1882 г., – это вид деления ядер (и клеток), приводящего к образованию из одной материнской клетки четырех дочерних клеток с уменьшением в два раза количества хромосом (рис. 16). В результате мейоза образуются гаплоидные клетки, имеющие одиночный набор хромосом. При мейозе происходит одна репликация ДНК, за которой следуют два митотических деления ядер и клеток (мейоз I и II).
Перед началом мейоза в интерфазе ДНК материнской клетки удваивается, но хромосомы остаются связанными своими центромерами, так что в ядре имеется по четыре набора каждой хромосомы; кроме того, в интерфазе увеличивается масса клетки и количество ее органелл. В каждом делении мейоза выделяются те же фазы, что и в митозе.
В мейозе I наиболее длительна профаза, во время которой происходит важнейшее событие кроссинговер (от англ. crossing-over – «перекрест») – перекрест гомологичных участков гомологичных хромосом с их последующим разрывом и присоединением участков хроматид к другой гомологичной хромосоме (рис. 17).

Рис. 16. Основные стадии мейоза (по Мамонтову)
А – протофаза I; Б – метафаза I; В – анафаза I; Г – телофаза I; Д – профаза II; Е – метафаза II, Ж – анафаза II, З – телофаза II

Рис. 17. Схема обмена генами двух хромосом (кроссинговер) (по Дж. Тортора и С. Грабовски)
Кроссинговер обеспечивает различные генетические комбинации. До кроссинговера каждая хромосома была либо материнской, либо отцовской, после него каждая хромосома содержит гены, происходящие как из отцовской, так и из материнской хромосомы, т. е. происходит генетическая рекомбинация.
Метафаза I напоминает аналогичную стадию митоза, хромосомы устанавливаются в экваториальной плоскости. В анафазе I гомологичные хромосомы отделяются друг от друга и расходятся к полюсам. В телофазе I наборы гомологичных хромосом находятся у полюсов, хотя их число уменьшилось вдвое, но каждая из них состоит уже из двух генетически различных хроматид. Формируются ядерная оболочка и ядрышко, образуются борозда деления, которая углубляется, и две клетки, каждая из которых содержит гаплоидный набор удвоенных хромосом, полностью разделяются.
Интерфаза II очень короткая, и, что самое главное, в ней не происходит удвоение ДНК. Фазы мейоза II не отличаются от описанных стадий митоза. Важным отличием является то, что в профазе II, которая происходит очень быстро, клетка содержит одиночный (гаплоидный) набор удвоенных хромосом, т. е. в каждой дочерней клетке по 46 хромосом. В результате мейоза II образуются четыре клетки, каждая из которых несет одиночный (гаплоидный) набор хромосом.
При мейотическом делении из каждой предшественницы половых клеток у женщин образуются одна яйцеклетка и три полярных тельца, которые рассасываются; у мужчин четыре сперматозоида. Во время образования половых клеток благодаря кроссинговеру создается множество различных сочетаний генов. При оплодотворении яйцеклетки сперматозоидом в зиготе восстанавливается диплоидный набор хромосом.
В зависимости от того, как распределился генетический материал во время образования яйцеклетки и сперматозоида, число возможных сочетаний генов в оплодотворенной яйцеклетке огромно. Вот почему каждый человек уникален. Оплодотворение приводит к тому, что каждый ген в зиготе представлен двумя экземплярами (аллелями) от отца и матери. Поэтому физические и психические особенности будущего ребенка будут зависеть от взаимодействия этих генов.
Цитозоль. Рибосомы и синтез белка
Цитозоль, представляющий собой часть цитоплазмы, окружающей органеллы, занимает 53–55 % общего объема клетки. В цитозоле содержится огромное количество ферментов, катализирующих различные реакции промежуточного обмена, а также белки цитоскелета. На расположенных в цитозоле рибосомах синтезируются многие белки. Рибосомы, осуществляющие синтез белка, присутствуют во всех клетках человека, кроме зрелых эритроцитов. Рибосомы расположены поодиночке (монорибосомы) или группами в виде розеток, спиралей, завитков (полирибосомы, или полисомы). Рибосомы и полисомы могут свободно располагаться в цитоплазме, или, это называлось ранее, прикрепляться к мембранам гранулярного эндоплазматического ретикулума. Свободные рибосомы синтезируют белок, необходимый для жизнедеятельности самой клетки, прикрепленные белок, подлежащий выведению из клетки.

Рис. 18. Общая схема белкового синтеза в клетке
Синтез белка (трансляция) связан с процессом транскрипции переписывания информации, хранящейся в ДНК. Информация о структуре белка, заключенная в ДНК, «переписывается» на информационную, или мессенджер (от англ. messenger – «посредник») РНК (мРНК).
Триплетный генетический код, расшифрованный в 60-х годах М. Ниренбергом, С. Очоа, X. Кораной, основан на триплетах, или кодонах три нуклеотида, определяют присоединение к полипептидной цепи одной аминокислоты. Небольшие транспортные РНК (тРНК) выполняют двойную функцию: они присоединяют молекулу аминокислоты, транспортируют ее в рибосому и узнают триплет, соответствующий этой аминокислоте в молекуле мРНК. Антикодон тРНК узнает кодон мРНК и спаривается с ним.
Реакции синтеза белка осуществляют рибосомы, которые считывают информацию, заложенную в мРНК, продвигаясь вдоль нее. Синтез белка начинается с того, что малая субъединица рибосомы связывается с инициаторной тРНК, несущей молекулу метионина. Этот комплекс присоединяется к инициаторному кодону мРНК, после этого к малой присоединяется большая субъединица рибосомы. К рибосоме подходит следующая тРНК и образуется первая пептидная связь. Перемещаясь по цепи мРНК, рибосома присоединяет следующие аминокислоты, которые связываются между собой, а молекулы тРНК отделяются, чтобы вскоре присоединить новую аминокислоту. При достижении рибосомой стоп-кодона синтез прекращается, и полипептидная цепь отделяется от рибосомы (рис. 18).
Ткани
Ткань – это исторически сложившаяся общность клеток и межклеточного вещества, объединенных единством происхождения, строения и функций. В организме человека выделяют четыре типа тканей: эпителиальные, соединительные, мышечные и нервную.
Эпителиальные ткани
Эпителиальные ткани покрывают поверхность тела и выстилают слизистые оболочки, отделяя организм от внешней среды (покровный эпителий), а также образуют железы (железистый эпителий). Кроме того, выделяют сенсорный эпителий, клетки которого изменены для восприятия специфических раздражений в органах слуха, равновесия и вкуса.
В зависимости от отношения к базальной мембране[3] покровный эпителий подразделяют на однослойный, все клетки которого лежат на базальной мембране, однако не обязательно все достигают поверхности; и многослойный, в котором лишь клетки нижнего слоя лежат на базальной мембране. Однослойный эпителий, в свою очередь, подразделяется на однорядный (плоский, кубический, цилиндрический) и многорядный. В однорядном эпителии ядра всех клеток эпителиального пласта расположены на одном уровне, в многорядном – на различных уровнях. Клетки многорядного эпителия могут быть снабжены микроворсинками, стереоцилиями или ресничками. В зависимости от формы клеток и их ороговения различают многослойный плоский неороговевающий эпителий и аналогичный ороговевающий эпителий, а также редко встречающийся у человека многослойный кубический и многослойный призматический столбчатый эпителий (рис. 19).
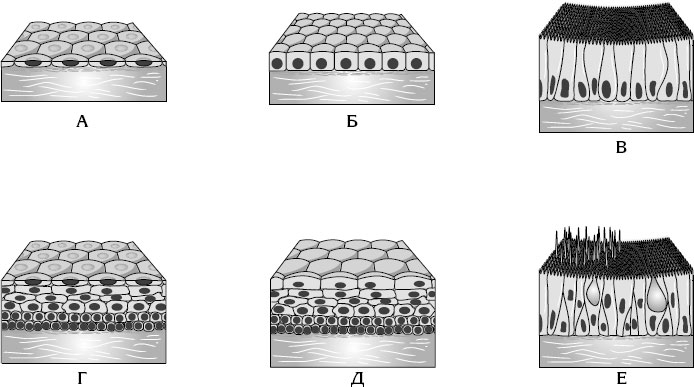
Рис. 19. Строение эпителиальной ткани
А – однослойный плоский эпителий; Б – однослойный кубический эпителий; В – однослойный цилиндрический эпителий Г – многослойный плоский неороговевающий эпителий; Д – переходный эпителий; Е – псевдомногослойный реснитчатый эпителий
Клетки эпителия (эпителиоциты), отличаются большим разнообразием форм и размеров. В зависимости от формы клеток различают следующие виды эпителиоцитов: плоские (эндотелиоциты и мезотелиоциты), кубические, столбчатые (микроворсинчатые, реснитчатые). Кроме того, имеются пигментированные эпителиоциты. Железистый эпителий состоит из железистых эпителиоцитов (гладулоцитов).
Строение клеток различных видов эпителия неодинаковы. Однако все они имеют и общие структурные особенности. Эпителиоциты полярны, их апикальные части отличаются от базальных; за редким исключением они образуют пласт, который располагается на базальной мембране и лишен кровеносных сосудов. В клетках представлены все описанные выше органеллы общего назначения, их развитие зависит от выполняемой клеткой функции. Так, клетки, секретирующие белок, богаты элементами гранулярного эндоплазматического ретикулума. Клетки, продуцирующие стероиды, – элементами гладкого эндоплазматического ретикулума, и в тех и в других хорошо развит комплекс Гольджи. Всасывающие клетки покрыты множеством микроворсинок, а эпителиоциты, покрывающие слизистую оболочку дыхательных путей, – ресничками.
Железа представляет собой орган, паренхима которого сформирована из высокодифференцированных железистых клеток (гландулоцитов). Железы подразделяются на экзокринные, имеющие выводные протоки; эндокринные, не имеющие выводных протоков и выделяющие синтезируемые ими продукты непосредственно в межклеточные пространства, откуда они поступают в кровь и лимфу; и смешанные, состоящие из экзо- и эндокринных отделов (например, поджелудочная железа) (рис. 20). В организме человека имеется множество одноклеточных желез бокаловидных клеток, лежащих среди других эпителиальных клеток, покрывающих слизистые оболочки полых органов пищеварительной, дыхательной и половой систем. Они вырабатывают слизь.
Экзокринная железа состоит из секреторного отдела, сформированного лежащими на базальной мембране железистыми клетками экзокриноцитами, которые вырабатывают различные секреты, и протоков. В зависимости от строения начального (секреторного) отдела различают трубчатые (напоминают трубку): ацинозные (напоминают грушу или удлиненную виноградину) и альвеолярные (напоминают шарик), а также трубчато-ацинозные и трубчато-альвеолярные железы, секреторные отделы которых имеют и ту и другую форму. В зависимости от строения протоков железы подразделяются на простые, имеющие один проток, и сложные, в главные выводные протоки которых вливается множество протоков, в каждый из них, в свою очередь, открывается несколько секреторных отделов. Железы вырабатывают различные секреты: белковый (серозные железы), слизь (слизистые) и смешанный.

Рис. 20. Схема строения экзокринных и эндокринных желез
А – экзокринная железа, Б – эндокринная железа; 1 – начальный отдел; 2 – секреторные гранулы; 3 – выводной проток экзокринной железы; 4 – покровный эпителий; 5 – соединительная ткань; 6 – кровеносный сосуд
Соединительные и опорные ткани
Соединительные ткани представляют обширную группу, включающую собственно соединительные ткани (рыхлая волокнистая и плотная волокнистая неоформленная и оформленная), ткани со специальными свойствами (ретикулярная, пигментная, жировая), твердые скелетные (костная, хрящевая) и жидкие (кровь и лимфа). Соединительные ткани выполняют опорную, механическую (плотная волокнистая соединительная ткань, хрящ, кость), трофическую (питательную), защитную (фагоцитоз и выработка антител) функции (рыхлая волокнистая и ретикулярная соединительная ткань, кровь и лимфа). В отличие от других тканей соединительные сформированы из многочисленных клеток и межклеточного вещества, состоящего из гликозаминогликанов, часть которых, связываясь с белками, образует протеогликаны, и различных волокон (коллагеновых, эластических, ретикулярных). Межклеточное вещество кости твердое, крови и лимфы – жидкое. Учитывая, что многие клетки крови являются одновременно и клетками соединительной ткани, а другие – их предшественниками, считаем целесообразным начать описание соединительных тканей с крови.
Кровь
Кровь состоит из клеток, взвешенных в жидком межклеточном веществе сложного состава (плазма). Кровь выполняет следующие функции: транспортную, трофическую (питательную), защитную, гемостатическую (кровоостанавливающую). Кроме того, кровь участвует в сохранении постоянного состава и свойств внутренней среды организма гомеостаза (от греч. homoios – «одинаковый» и stasis – «состояние, неподвижность»). Общее количество крови у взрослого человека 4–6 л, что составляет 6–8 % массы его тела (у мужчин в среднем около 5,4 л, у женщин – около 4,5 л).
Плазма – это жидкая часть крови, в которой содержится до 91 % воды, 6,5–8,0 % белков, около 2 % низкомолекулярных соединений, рН плазмы колеблется в пределах от 7,37 до 7,43, а удельный вес 1,025–1,029. Плазма богата как электролитами, так и неэлектролитами.
Белки плазмы крови (6,5–8,0 г/л, альбумины и глобулины) выполняют трофическую, транспортную, защитную, буферную функции; они также участвуют в свертывании крови и создании коллоидно-осмотического давления. На долю плазмы приходится около 54 % объема крови, на долю форменных элементов – около 44 %. В крови содержатся безъядерные клетки эритроциты (4,0–5,0) × 1012/л, лейкоциты (4,0–6,0) × 109/л, среди которых выделяют зернистые, или гранулоциты (нейтрофильные, ацидофильные и базофильные), а также незернистые, или агранулоциты (моноциты). В крови имеются также кровяные пластинки (тромбоциты), число которых составляет (180,0–320,0) × 109 в литре и лимфоциты, являющиеся структурными элементами лимфоидной системы (рис. 21).

Рис. 21. Кровь
А – мазок периферической крови взрослого человека (общий вид): 1 – эритроциты; 2 – лимфоциты; 3 – моноцит; 4 – нейтрофильные гранулоциты; 5 – эозинофильные гранулоциты; 6 – базофильные гранулоциты; 7 – тромбоциты; Б – клетки крови: I – базофильный гранулоцит; II – ацидофильный гранулоцит; III – сегментоядерный нейтрофильный гранулоцит; IV – эритроцит; V – моноцит; VI – тромбоциты; VII – лимфоцит
Эритроциты (от греч. Erythros – «красный»), или красные кровяные тельца, имеют форму двояковогнутых дисков диаметром от 7 до 10 мкм, они содержат гемоглобин, осуществляющий перенос О2 и СО2. Содержание эритроцитов составляет: м. 4 × 1012–5,6 × 1012/л, ж. 3,4 × 1012–5,0 × 1012/л. Общее количество эритроцитов у мужчин достигает 25 × 1012, у женщин – 18 × 1012 клеток, а общая площадь поверхности всех эритроцитов около 3800 м2. Эритроцит – единственная клетка в теле человека, которая не содержит ядра. Клетка покрыта плазмолеммой толщиной около 7 нм, в которую встроены антигены систем АВО и резус (группы крови и резус-фактор), мембранные ферменты. Длительность жизни эритроцитов – около 120 дней, после чего они разрушаются и поглощаются макрофагоцитами в селезенке, костном мозге и печени (рис. 22).

Рис. 22. Нормальные эритроциты в форме двояковогнутого диска
Лейкоциты (от греч. leukos – «белый») представляют собой ядросодержащие клетки, обладающие амебоидной подвижностью. В отличие от эритроцитов, которые выполняют присущие им функции в просвете кровеносных сосудов, лейкоциты осуществляют свои в тканях, куда они мигрируют посредством диапедеза (от греч. dia – «сквозь», pedesis – «прыжок»). Содержание лейкоцитов составляет: у мужчин – 4,3 × 109–11,3 × 109/л, у женщин – 3,2 × 109–10,2 × 109/л.
К зернистым лейкоцитам (гранулоцитам) относятся нейтрофильные, или полиморфноядерные, которые составляют от 47 до 72 % всех лейкоцитов, содержание их составляет 2,0–5,5 × 109/л, 2000–5500 в 1 мкл крови, а общее их количество в крови взрослого человека колеблется в пределах 3 × 1012. Время циркуляции в крови не превышает 8–12 ч, затем посредством диапедеза они мигрируют в соединительную ткань. Зрелый нейтрофильный гранулоцит представляет собой сферическую клетку диаметром 10–12 мкм с дольчатым ядром. В ядрах нейтрофильных гранулоцитов женщин имеются тельца полового хроматина (тельца Барра)[4] диаметром до 1,5–2,0 мкм. Цитоплазма гранулоцита богата гранулами двух типов: нейтрофильными и азурофильными. Мелкие преобладающие специфические нейтрофильные гранулы в световом микроскопе выглядят лиловыми. Они богаты щелочной фосфатазой и бактерицидным веществом. Более крупные красновато-фиолетовые азурофильные гранулы составляют 20–30 % всех гранул. Оба типа гранул участвуют в фагоцитозе и инактивации фагоцитированного материала (рис. 23).
Осуществляя фагоцитоз продуктов распада и микроорганизмов, нейтрофильные гранулоциты погибают, а освобождающиеся при этом лизосомальные ферменты разрушают окружающие ткани, способствуя формированию гнойника. В состав гноя обычно входят разрушенные нейтрофильные гранулоциты и продукты распада ткани. Количество нейтрофильных гранулоцитов резко возрастает при острых воспалительных и инфекционных заболеваниях.
Эозинофильные (ацидофильные) гранулоциты составляют 0,5–5,0 % циркулирующих лейкоцитов. В 1 мкл крови их число колеблется в пределах от 20 до 300 (0,02–0,3 × 109/л). Они циркулируют в крови не более 8 дней, после чего покидают кровеносное русло через мелкие венулы и проникают в рыхлую соединительную ткань. Особенно много их в слизистой оболочке кишечника и дыхательных путей. Диаметр эозинофильных гранулоцитов 10–15 мкм. Их двухлопастное ядро напоминает по форме гантель. В цитоплазме имеется множество крупных эозинофильных (красных или оранжевых) светопреломляющих несколько удлиненных гранул, которые представляют собой лизосомы. Эозинофильные гранулоциты осуществляют фагоцитоз, однако менее активно, чем нейтрофильные. Их подвижность также менее выражена. Эозинофильные гранулоциты участвуют в иммунных реакциях. Количество эозинофильных гранулоцитов в циркулирующей крови (эозинофилия) увеличивается при паразитарных заболеваниях, аллергических и аутоиммунных процессах.
Количество базофильных гранулоцитов в циркулирующей крови невелико – около 0,5 % всех лейкоцитов (0–60 клеток в 1 мкл крови, 0–0,65 × 109/л), а время их циркуляции не превышает 12–15 ч. Диаметр клетки 10–12 мкм, в световом микроскопе в клетке видно множество крупных темно-синих округлых или овальных гранул. Количество их столь велико, что они маскируют крупное ядро. Гранулы содержат гистамин и гепарин. Базофильные гранулоциты также осуществляют фагоцитоз и участвуют в аллергических реакциях.

Рис. 23. Гранулоциты (по Юриной и Румянцевой)
I – сегментоядерный нейтрофильный гранулоцит; 1 – вторичные (специфические) гранулы; 2 – первичные (азурофильные) гранулы; 3 – сегменты ядра; 4 – периферическая зона, не содержащая органелл; 5 – тельце полового хроматина; 6 – микроворсинки и псевдоподии; 7 – вторичные (специфические) гранулы; II – ‘озинофильный (ацидофильный) гранулоцит 8 – зрелые специфические гранулы, содержащие кристаллоиды; 9 – сегменты ядра; 10 – первичные (азурофильные) гранулы; III – базофильный гранулоцит; 11 – сегменты ядра; 12 – гранулы базофила различной величины и плотности
В крови постоянно присутствуют также лимфоциты, которые являются структурными элементами лимфоидной (иммунной) системы. Лимфоциты содержатся в большом количестве в крови (19–37 % всех лейкоцитов, 1200–3000 в 1 мкл, 1,2–3,0 × 109/л), преобладают в лимфе и ответственны за иммунитет. В организме взрослого человека их число достигает 6–1012. Все лимфоциты имеют сферическую форму, но отличаются друг от друга своими размерами. Диаметр большей части лимфоцитов около 8 мкм (малые лимфоциты). Лимфоциты подразделяются на две категории: тимусзависимые (Т-лимфоциты), которые осуществляют в основном клеточный иммунитет, и бурсозависимые (В-лимфоциты), осуществляющие гуморальный иммунитет. Морфологически они не отличаются друг от друга.
Моноциты составляют от 3 до 11 % циркулирующих лейкоцитов крови (90–600 в 1 мкл, 0,09–0,6 × 109/л). Время их пребывания в кровеносной системе 2–3 дня, после чего они мигрируют в ткани, где превращаются в макрофаги и выполняют свою главную функцию защиту организма. Моноцит – клетка овальной формы, диаметром около 15 мкм. Крупное почкообразное богатое хроматином ядро окружено большим количеством голубоватой цитоплазмы, в которой присутствуют мелкие азурофильные гранулы (первичные лизосомы). В клетке имеется умеренное количество органелл (рис. 24).

Рис. 24. Моноцит (по Крстичу, с изм.)
1 – лизосома; 2 – митохондрия; 3 – ядро; 4 – комплекс Гольджи
Тромбоциты, или кровяные пластинки, уплощенные овальные двояковыпуклые безъядерные фрагменты крупных клеток мегакриоцитов диаметром 2–4 и толщиной 0,5–0,75 мкм. Количество их достигает 180–320 тыс. в 1 мкл крови (180,0 × 109–320,0 × 109/л). Время их циркуляции в крови не превышает семи дней, после чего они попадают в селезенку и легкие, где разрушаются. Тромбоциты участвуют в свертывании крови, остановке кровотечений и в защите организма благодаря способности фагоцитировать вирусы, иммунные комплексы и неорганические частички. При повреждении стенок мелких кровеносных сосудов кровотечение прекращается в течение 1–3 мин (первичный гематиз), при ранении более крупного кровеносного сосуда тромбоциты прилипают к ним и реагируют, в результате чего из них высвобождаются биологически активные вещества, которые вызывают сужение сосудов. Под действием одного из них белок плазмы протромбин, образующийся в печени, превращается в тромбин, который вызывает переход плазменного белка фибриногена, также образующегося в печени, в фибрин. Последний и формирует основную часть тромба (рис. 25).

Рис. 25. Тромбоцит (по Альбертсу и соавт., с изм.)
1 – пучок микротрубочек; 2 – грануломер; 3 – гиаломер
Собственно соединительные ткани
Рыхлая волокнистая соединительная ткань (РВСТ) располагается преимущественно по ходу кровеносных и лимфатических сосудов, нервов, покрывает мышцы, образует строму (каркас) большинства внутренних органов, собственную пластинку слизистой оболочки, подсерозную основу, адвентициальную оболочку. РВСТ состоит из многочисленных собственных и пришлых клеток (фибробласты, фиброциты, ретикулярные, перициты, макрофагоциты, тканевые базофилы, плазмоциты, жировые, пигментные, лимфоциты, гранулярные лейкоциты), которые располагаются в межклеточном веществе (рис. 26). Последнее продуцируется в основном фибробластами и представлено коллагеновыми, эластическими, ретикулярными волокнами и основным веществом. По мере старения фибробласты превращаются в многоотростчатые фиброциты, которые образуют трехмерную сеть, в пространствах которой располагаются различные клетки. Тучные клетки лежат вблизи капилляров, перициты окружают капилляры, будучи внедрены в их базальную мембрану. Наряду с клетками, синтезирующими компоненты межклеточного вещества, в РВСТ присутствуют и клетки, разрушающие его фиброкласты, которые по своей структуре весьма напоминают фибробласты; они фагоцитируют коллаген и богаты лизосомами.
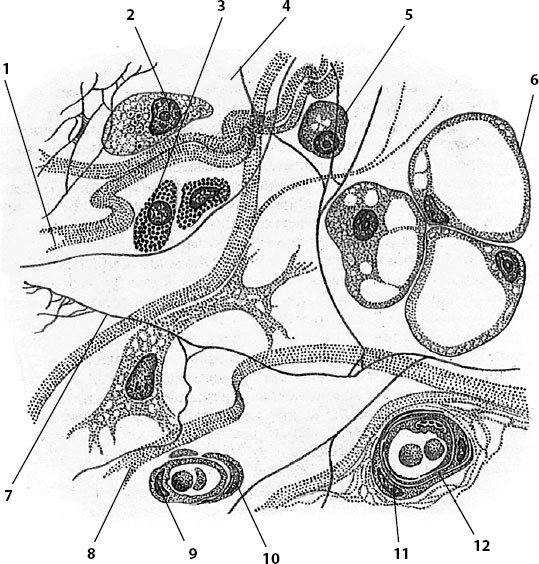
Рис. 26. Рыхлая волокнистая соединительная ткань
1 – коллагеновое волокно; 2 – макрофагоцит; 3 – тканевый базофил; 4 – аморфное межклеточное (основное) вещество; 5 – плазмоцит (плазматическая клетка); 6 – адипоцит (жировая клетка); 7 – эластическое волокно; 8 – фибробласт; 9 – эндотелиоцит; 10 – перицит; 11 – миоцит; 12 – кровеносный сосуд
Коллаген образован тремя полипептидными α-цепями, которые, скручиваясь, образуют молекулу тропоколлагена. Коллаген имеет спиральное строение, что обеспечивает создание весьма прочных малорастяжимых структур. Коллагеновые волокна толщиной 1–20 мкм – это изогнутые тяжи, состоящие из фибрилл с поперечной исчерченностью. Эластические волокна толщиной от 3 до 10 мкм образованы белком эластином, который также синтезируется фибробластами. В отличие от коллагеновых эластические волокна способны растягиваться в 1,5 раза, после чего возвращаются в исходное состояние. Эластические волокна анастомозируют и переплетаются между собой, образуя сети, окончатые пластины и мембраны. Тонкие (от 100 нм до 1,0 мкм), разветвленные, малорастяжимые ретикулярные волокна, переплетаясь между собой, образуют мелкопетлистую сеть, в ячейках которой расположены клетки. Ретикулярные волокна образуют каркасы лимфоидных органов (органов кроветворения и иммунной системы), печени, поджелудочной железы и других паренхиматозных органов, окружают капилляры, кровеносные и лимфатические сосуды, а также связаны с ретикулярными клетками.

Рис. 27. Макрофаг (по Крстичу, с изм.)
1 – ядро; 2 – фаголизосомы (вторичные лизосомы); 3 – псевдоподии; 4 – митохондрия; 5 – остаточное тельце; 6 – цистерны гранулярной эндоплазматической сети
Макрофаг (макрофагоцит). В 1882 г. И. И. Мечников впервые описал фагоцитоз. За это открытие он был удостоен Нобелевской премии. После двух-трех суток циркуляции моноцит покидает кровеносное русло. За сутки у человека обменивается 0,5–1,0 × 109 моноцитов, причем количество их в циркулирующей крови примерно в 20 раз меньше, чем в тканях. В 70-х годах XX века сформировалось представление о системе мононуклеарных фагоцитов (СМФ), включающей группу клеток, объединенных общностью происхождения из моноцитов крови (которые, в свою очередь, имеют общего предшественника стволовую кроветворную клетку), строения и функции – активный фагоцитоз и пиноцитоз (рис. 27).
Основной особенностью макрофагов является большое количество различных функциональных форм лизосом. Макрофаги секретируют большое количество различных биологически активных веществ. Зрелые макрофаги не способны делиться. В то же время СМФ является саморегулируемой системой. В ответ на раздражение периферического звена (тканевые макрофаги), усиливается деление (пролиферация) клеток-предшественниц моноцитов в костном мозге, увеличивается количество моноцитов костного мозга и крови, они мигрируют в ткани и превращение в макрофаги.
ВНИМАНИЕ
Основные функции макрофагов – это участие в естественном, специфическом, противоопухолевом иммунитете и секреция различных биологически активных веществ.
Плазматические клетки (пл. кл.) вырабатывают антитела (иммуноглобулины), это определяет их важнейшую роль в защите организма. Следует обратить внимание на происхождение пл. кл. из В-лимфоцитов. Плазмоциты – белоксинтезирующие клетки, богатые элементами зернистого ЭР, располагающиеся вблизи мелких кровеносных сосудов в лимфоидных органах, в слизистой оболочке пищеварительной и дыхательной систем и т. д. (рис. 29).
Тучные клетки содержат множество крупных (до 2 мкм) мембранных гранул, богатых биологически активными веществами, влияющими на кровеносные сосуды.

Рис. 28. Происхождение и структура системы мононуклеарных фагоцитов (СМФ)
Ретикулярная клетка (Р. кл.) – удлиненная клетка, ее многочисленные отростки соединяются с отростками других Р. кл., формируя сеть. Ретикулярные волокна располагаются в углублениях, образованных плазмалеммой клетки и ограниченных ею. При неблагоприятных условиях (инфекция, внедрение инородных частиц и т. д.) Р. кл. округляются, отделяются от ретикулярных волокон и становятся способными к фагоцитозу. Ретикулярные клетки образуют строму органов иммунной системы и кроветворения (рис. 30