Глава 2. Молекулярные основы жизни
Фундаментальную роль в структуре живой материи играет первый уровень ее организации – молекулярный. На этом уровне анализируются химические вещества, составляющие живой организм, взаимосвязь их структуры и функции.
Все имеющиеся в клетке биохимические соединения можно разделить на неорганические (вода и минеральные соли) и органические (соединения углерода).
2.1. Органические соединения в составе живых организмов
Органические соединения характерны только для живых организмов. Можно сказать, что жизнь на Земле построена на основе углерода, который обладает рядом уникальных свойств. Основное значение для выполнения роли «кирпичиков» жизни имеет способность атомов углерода соединяться друг с другом, образуя цепи и кольца. Именно этой способностью и объясняется чрезвычайное разнообразие органических соединений.
Разнообразие органических соединений
Хотя органические молекулы составляют менее 1 % всех молекул клетки (99 % молекул приходится на долю воды), они определяют протекание основных биохимических процессов. В клетке встречаются как малые органические соединения (содержащие до 20 или чуть более атомов углерода), так и большие, сложные молекулы – биополимеры, состоящие из многих единиц – мономеров.
Биополимеры образуют 4 класса биомолекул: белки, нуклеиновые кислоты, углеводы и липиды. Ключевую роль в жизнедеятельности организмов играют белки и нуклеиновые кислоты. Углеводы и липиды представляют собой очень разнородные группы, объединяющие соединения с различной структурой и биологическими функциями. Деление на эти группы – скорее дань исторической традиции, чем результат биохимического анализа. С них мы начнем рассмотрение молекулярного уровня организации жизни.
Углеводы
Углеводы – это наиболее распространенная в природе группа органических веществ. Основная их функция – энергетическая. Все углеводы содержат гидроксильные группы (—ОН) вместе с альдегидной или кетогруппой. Выделяют три группы углеводов (табл. 2.1).
Наибольшее значение в природе среди моносахаридов имеют пентозы (рибоза, дезоксирибоза, рибулоза) и гексозы (глюкоза, фруктоза, галактоза). Производными моносахаридов являются «сахарные» кислоты (к ним относятся, например, аскорбиновая кислота – витамин С), многоатомные спирты, гликозиды (к ним относятся некоторые антибиотики – эритромицин, стрептомицин, пуромицин и др.), аминосахара. Все их можно рассматривать как моносахара с дополнительной функциональной группой (—СООН, – NH2 и др.).
Среди олигосахаридов наиболее значимы мальтоза (глюкоза + глюкоза), сахароза (глюкоза + фруктоза), лактоза (глюкоза + галактоза), рафиноза (глюкоза + фруктоза + галактоза). Чрезвычайно важна роль гликопротеидов, определяющих сигналы узнавания на клеточном уровне. Нарушения «сигнализации» приводят к многочисленным патологиям, в том числе и к злокачественным новообразованиям.
Таблица 2.1. Классификация углеводов

К важнейшим гомополисахаридам относятся целлюлоза, гликоген, крахмал. Мономерами их является глюкоза, а различия определяются особенностями структуры (линейной или разветвленной).
Гетерополисахариды обычно состоят из повторяющихся дисахаридов. Примерами их могут служить хитин и муреин, выполняющие структурную функцию в клетках грибов, бактерий, членистоногих, а также мукополисахариды – важнейший элемент соединительной ткани животных с разнообразными функциями.
Использование конкретных полисахаридов как структурных и энергетических компонентов клетки является одной из фундаментальных характеристик при делении представителей живой природы на отдельные царства.
Липиды
Липиды – это обширная группа неполярных, нерастворимых в воде органических соединений. Они отличаются большим разнообразием, но в общем виде представляют собой сложные эфиры какого-либо спирта и жирной кислоты.
Жирные кислоты – это карбоновые кислоты с длинной (12–20 атомов углерода) цепью. Они могут быть насыщенными (содержат только С – С-связи) или ненасыщенными (содержат одну или несколько С = С-связей).
Липиды делятся на простые и сложные (табл. 2.2).
Наиболее распространены в природе жиры (как универсальный источник энергии) и фосфолипиды (как неотъемлемый компонент клеточной мембраны). Важную защитную функцию выполняют воски. Гликолипиды являются компонентами миелиновой оболочки нервных волокон.
Особую группу веществ составляют производные липидов. Наибольшее значение среди них имеют полициклические соединения – стероиды (к ним относятся холестерин, стероидные гормоны, желчные кислоты) и пигменты каротиноиды. Производными липидов являются все жирорастворимые витамины (А, D, Е, К).
Таблица 2.2. Классификация липидов

Белки
Белки имеют первостепенное значение в жизни организмов. Огромное разнообразие живых существ в значительной степени определяется различиями в составе имеющихся в их организме белков. Например, в организме человека их известно более 5 млн.
Белки – это полимеры, мономерами которых являются аминокислоты (рис. 2.1).

Рис. 2.1. Структура аминокислоты
В природных белках встречается 20 различных аминокислот, которые отличаются друг от друга только радикалами (R). Все они являются α-аминокислотами, так как карбоксильная группа и аминогруппа у них присоединена к одному атому α-углерода.
Кроме этих двадцати, в природных организмах известно около 300 других аминокислот (примерно 150 аминокислот имеется у человека), находящихся в разных клетках в свободном состоянии или в составе низкомолекулярных соединений. Некоторые из них играют исключительно важную роль.
Аминокислоты способны вступать в реакцию поликонденсации и формировать полипептидную цепочку. Образующаяся при этом —CO – NH-связь называется пептидной связью.
Условно к белкам относят полипептиды, состоящие из 50 или более аминокислотных остатков. При меньшем числе аминокислот вещества называются пептидами. Многие гормоны и антибиотики являются пептидами, состоящими из 2–20 аминокислотных остатков.
В молекуле белка может содержаться до нескольких тысяч аминокислот. Специфичность белков определяется набором используемых аминокислот, последовательностью их расположения в полипептидной цепочке и специфичностью укладки этой цепочки. Некоторые белки состоят из нескольких полипептидных цепей, соединенных вместе. Часто такие полипептидные цепи удерживаются небелковым компонентом. Наличие небелкового компонента характерно для группы сложных белков (гликопротеины, нуклеопротеины, липопротеины, гемоглобины и др.). Простые белки состоят только из полипептидов.
Белки в организме выполняют чрезвычайно многообразные функции. Большинство важнейших биохимических процессов осуществляется белками. Они играют ключевую роль в реализации процессов дыхания, движения, иммунитета, входят в состав большинства клеточных и тканевых структур.
Особо следует отметить каталитическую роль белков, поскольку почти все ферменты (биологические катализаторы) являются белками. Каждый фермент катализирует только одну реакцию или действует только на один тип связи. Обычно ферменты представляют собой сложные белки с небелковой частью – коферментом. Многие витамины являются предшественниками коферментов, в чем и заключается их биологическое значение.
2.2. Нуклеиновые кислоты
Нуклеиновые кислоты обеспечивают хранение и воспроизведение наследственной информации. Этим определяется их фундаментальное значение для жизни на Земле.
Нуклеиновые кислоты – это полимеры, мономерами которых являются нуклеотиды. Нуклеотид включает в себя азотистое основание, углевод и остаток фосфорной кислоты (рис. 2.2).

Рис. 2.2. Структура нуклеотида
В природе встречаются два вида нуклеиновых кислот: ДНК и РНК.
Дезоксирибонуклеиновая кислота
Дезоксирибонуклеиновая кислота (ДНК) является местом хранения генетической информации организмов, поэтому можно сказать, что это «самая главная молекула». Молекула ДНК состоит из двух полинуклеотидных цепей, спирально закрученных одна относительно другой.
В составе нуклеотидов ДНК встречаются 4 типа азотистых оснований: А – аденин; Т – тимин; Г – гуанин; Ц – цитозин.
Углевод – дезоксирибоза.
В полинуклеотидной цепочке соседние нуклеотиды связаны между собой ковалентными связями между дезоксирибозой одного и остатком фосфорной кислоты другого нуклеотида. Две полипептидные цепочки объединяются в единую молекулу ДНК при помощи водородных связей между азотистыми основаниями нуклеотидов разных цепей. Соединены азотистые основания по принципу комплементарности: аденин с тимином, гуанин с цитозином.
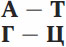
Принцип комплементарности – это один из фундаментальных законов природы, определяющий механизм передачи наследственной информации.
Хотя в молекуле ДНК всего 4 типа разных нуклеотидов, благодаря различной их последовательности и огромному числу (до нескольких десятков тысяч) в полинуклеотидной цепочке достигается невероятное разнообразие этих молекул.
Рибонуклеиновая кислота
Молекулы рибонуклеиновой кислоты (РНК), несмотря на их разнообразие, построены по общим структурным принципам. Они состоят из одной полинуклеотидной цепочки, значительно более короткой, чем цепочка ДНК. В нуклеотидах имеются 4 типа азотистых оснований, из которых специфичным для РНК является урацил: А – аденин; У – урацил; Г – гуанин; Ц – цитозин.
Углевод – рибоза.
В клетке имеется несколько типов РНК, из которых основными являются три.
Информационная (и-РНК), или матричная (м-РНК). Переносит информацию о структуре белка с ДНК на рибосомы – место непосредственного синтеза полипептидной цепочки. Каждый белок клетки кодируется своей и-РНК.
Рибосомальная (р-РНК). Входит в состав рибосом. Помимо структурной функции, принимает непосредственное участие в синтезе полипептидной цепочки. Этот тип составляет 85 % всей РНК клетки.
Транспортная (т-РНК). Переносит аминокислоты к месту синтеза белков на рибосомы. Каждой аминокислоте соответствует своя т-РНК, которая имеет специфическую петлевую структуру (форма «клеверного листа»). Центральная петля молекулы содержит антикодон – триплет нуклеотидов, определяющий специфичность данной т-РНК и ее соответствие определенной аминокислоте.
Свободные нуклеотиды
Не только нуклеиновые кислоты, но и свободные нуклеотиды и их производные играют важную роль в жизни клетки. Центральную роль в энергетическом обмене клетки играет аденозинтрифосфорная кислота (АТФ), имеющая три остатка фосфорной кислоты (рис. 2.3). Все виды активности организмов (движение, синтез, деление и др.) происходят за счет энергии, высвобождаемой при гидролизе АТФ. Эта энергия запасается в высокоэнергетических (макроэргических) связях между остатками фосфорной кислоты.
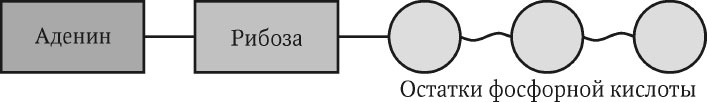
Рис. 2.3. Структура аденозинтрифосфорной кислоты
Гидролиз любой из двух макроэргических связей приводит к выделению энергии. АТФ чрезвычайно быстро обновляется. У человека каждая молекула АТФ расщепляется и вновь восстанавливается более 2400 раз в сутки.
Из нуклеотидов построены и некоторые другие важнейшие молекулы клетки: циклический аденозинмонофосфат (цАМФ) – одна из основных сигнальных молекул клетки, кофермент А (СоА), играющий ключевую роль в обмене веществ, никотинамидадениндинуклеотид (НАД) и флавинадениндинуклеотид (ФАД) – универсальные переносчики электронов и др. Структуру НАД, ФАД и СоА можно представить в общем виде (рис. 2.4). Предшественниками радикала у этих соединений являются водорастворимые витамины (РР, В2, В3), что объясняет их незаменимость в организме.
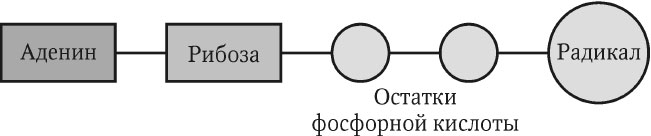
Рис. 2.4. Общая структура некоторых нуклеотидсодержащих молекул
2.3. Обмен веществ и энергии
Вся совокупность химических реакций, протекающих в живых организмах, называется обменом веществ, или метаболизмом. В результате этих реакций энергия, запасенная в химических связях, переходит в другие формы, т. е. обмен веществ всегда сопровождается обменом энергии. Первичным источником энергии для всего живого на Земле служит Солнце.
Многие организмы имеют уникальные метаболические пути, однако наиболее поразительно то общее, что присуще процессам метаболизма в живой природе. Несмотря на величайшее разнообразие живых организмов, отчетливо проявляется единство этих процессов. Выделяют две группы процессов метаболизма.
Анаболизм (ассимиляция) – совокупность процессов синтеза, идущих с потреблением энергии.
Катаболизм (диссимиляция) – совокупность процессов распада, сопровождающихся выделением энергии.
Анаболизм и катаболизм самым тесным образом взаимосвязаны: катаболические реакции дают «сырье» и энергию для анаболических процессов, в которых эта энергия запасается.
Все живые организмы можно разделить на группы, в зависимости от типа ассимиляции (рис. 2.5).
Автотрофы – организмы, способные самостоятельно синтезировать органические вещества из неорганических.
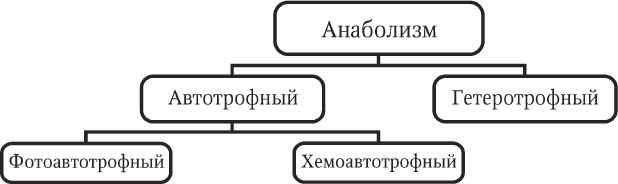
Рис. 2.5. Классификация типов анаболизма
Гетеротрофы – организмы, не способные синтезировать органические вещества из неорганических и нуждающиеся в поступлении готовых органических соединений.
Среди протистов можно выделить группу автогетеротрофных организмов, которые в зависимости от условий осуществляют либо автотрофный, либо гетеротрофный способ питания.
Фотоавтотрофы – организмы, использующие для синтеза энергию Солнца.
Хемоавтотрофы – организмы, использующие для синтеза энергию химических реакций.
Совокупность реакций катаболизма, протекающих во всех живых клетках, представляет собой разнообразные процессы биологического окисления. Поскольку запасенная в процессе ассимиляции энергия недоступна для непосредственного использования клеткой, основной функцией процесса биологического окисления является обеспечение организма энергией в доступной форме (прежде всего в виде АТФ). В природе организмы используют два пути получения энергии: аэробный распад (дыхание), проходящий в присутствии кислорода, и анаэробный распад (брожение), проходящий без кислорода (рис. 2.6). Соответственно организмы, реализующие эти пути, называются аэробами и анаэробами.

Рис. 2.6. Классификация типов катаболизма
2.4. Фотосинтез и хемосинтез
Основной путь, по которому практически вся энергия попадает в биосферу – это процесс фотосинтеза. Каждый год на Земле в процессе фотосинтеза образуется более 150 × 109 тонн органических веществ. Вклад хемоавтотрофов (к ним относятся только некоторые представители бактерий) в синтез органических соединений на Земле ничтожен по отношению к фотоавтотрофам.
Понятие фотосинтеза
Фотосинтез – это процесс преобразования энергии света в энергию химических связей органических соединений. У растений этот процесс локализован в особых клеточных органоидах – хлоропластах, а у фотосинтезирующих бактерий происходит в самой клетке.
Ключевую роль в процессе фотосинтеза играют фотосинтетические пигменты – вещества, способные поглощать свет. Выделяют три группы фотосинтетических пигментов: хлорофиллы (зеленые пигменты), каротиноиды (желтые и оранжевые пигменты), фикобилины (голубые и красные пигменты). В каждой группе имеется несколько подгрупп пигментов, поглощающих свет в разных областях спектра. Такое многообразие пигментов (только хлорофиллов известно более 10) обусловлено эволюционным приспособлением для обеспечения наибольшей эффективности поглощения солнечного света в различных условиях.
Пигментный состав является важным диагностическим признаком для систематики, поскольку он отражает филогенетические отношения организмов.
Основным пигментом, участвующим в фотохимических реакциях, является хлорофилл. В хлоропластах растений пигменты (по 200–400 молекул) собраны в функциональные единицы – фотосистемы. В фотосистемах молекула хлорофилла А выполняет роль реакционного центра, а молекулы всех других пигментов называются антенным комплексом, так как, подобно антеннам, собирают свет для реакционного центра и передают ему энергию квантов. Существуют два вида фотосистем.
Фотосистема I – реакционный центр образован молекулой хлорофилла А – Р-700.
Фотосистема II – реакционный центр образован молекулой хлорофилла А – Р-680.
Обычно обе фотосистемы работают синхронно, но фотосистема I может работать самостоятельно.
Стадии фотосинтеза
В процессе фотосинтеза различают две стадии – световую и темновую.
Во время световой фазы фотосинтеза энергия Солнца используется для синтеза АТФ и высокоэнергетических переносчиков электронов. Световая энергия, поглощенная любой молекулой пигмента, переносится на другую молекулу фотосистемы, пока не достигнет своего реакционного центра, после чего по специальной системе переносчиков возникает направленный поток электронов.
Существуют два варианта работы фотосистем во время световой фазы.
Нециклическое фотофосфорилирование – это основной путь, при котором обе фотосистемы работают синхронно.
Под действием света электроны реакционного центра фотосистемы II возбуждаются, переходят на акцептор электронов и переносятся по электронно-транспортной цепи к фотосистеме I. Потерянные электроны хлорофилла Р-680 заменяются электронами молекул воды в результате одновременного процесса фотолиза – ферментативного расщепления воды. В фотосистеме I электроны под действием света возбуждаются, переносятся на свой акцептор и через белок ферредоксин – на никотинамидадениндинуклеотидфосфат (НАДФ), который восстанавливается (НАДФ-Н2). Электроны хлорофилла Р-700 заменяются электронами фотосистемы II. Таким способом осуществляется однонаправленный поток электронов от реакционного центра фотосистемы II к НАДФ (рис. 2.7).
Циклическое фотофосфорилирование – это вариант световой фазы, когда возбужденные электроны фотосистемы I с акцептора направляются не на НАДФ, а возвращаются на цепь переносчиков фотосистемы II. При таком способе происходит образование АТФ, но не происходит образования НАДФ-Н2 (на рис. 2.7 обозначен пунктиром). Предполагается, что это более древний и примитивный вариант фотосинтеза.
В роли переносчиков электронов по электронно-транспортной цепи выступают различные сложные белки, содержащие гем, железо, медь (цитохромы, пластохинон, пластоцианин и др.). Процесс синтеза АТФ контролируется у растений особым ферментом АТФ-синтетазой.
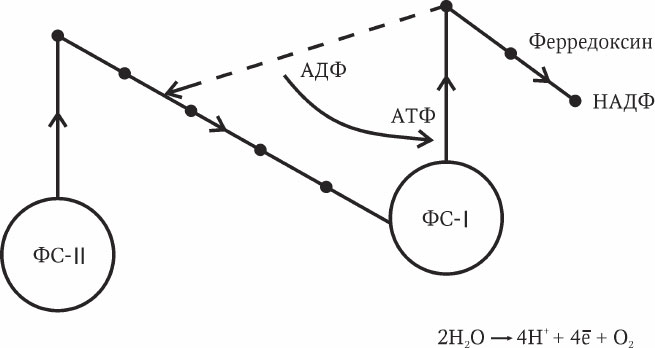
Рис. 2.7. Общая схема световой фазы фотосинтеза
В темновой фазе происходит превращение СО2 в углеводы, благодаря синтезированным в первой стадии АТФ и НАДФ. Существует несколько вариантов фиксации углекислоты.
Основной вариант – это цикл Кальвина, или С-3-путь, названный так потому, что непосредственный продукт фиксации СО2 представляет собой трехуглеродное соединение (глицерол-3-фосфат). Углекислота первоначально соединяется с 5-углеродным рибулозобифосфатом (РБФ), давая промежуточное 6-углеродное соединение. Цикл Кальвина включает в себя 13 последовательных реакций. На заключительном этапе вновь образуется РБФ, и цикл замыкается.
Шесть оборотов цикла дают одну молекулу глюкозы, для чего требуется 18 молекул АТФ и 12 молекул НАДФ, что можно выразить суммарным уравнением:
6СО2 →С6Н12О6.
Существует другой вариант синтеза углеводов при темновой фазе.
Это цикл Хэтча – Слэка, или С-4-путь, обнаруженный более чем у 100 растений жарких и засушливых районов. Этот путь требует больших энергозатрат, но имеет свои экологические преимущества.
Бактериальный фотосинтез и хемосинтез
Различные группы фотосинтезирующих бактерий осуществляют особую разновидность фотосинтеза – бактериальный фотосинтез. Бактерии имеют свои специфические фотосинтетические пигменты (в частности, различные бактериохлорофиллы), отличные от пигментов эукариот. Донорами Н+ у них служит не вода, а различные органические соединения (сероводород, спирты, жирные кислоты и др.), потому при бактериальном фотосинтезе не выделяется кислород. Рассмотрев все виды фотосинтеза, можно вывести общее уравнение этого процесса:
nСО2 + Н2А → (СН2О)n + H2 О + А,
где Н2А – обобщенный донор Н+.
Хемосинтез – это синтез органических веществ за счет энергии химических реакций. Он представляет собой другую форму автотрофной ассимиляции, свойственной некоторым бактериям. При хемосинтезе источником энергии служит не солнечный свет, а окисление неорганических соединений.
Нитрифицирующие бактерии окисляют образующийся при гниении органических остатков аммиак до нитрита, а затем до нитрата: NH3 → HNO2 →HNO3.
Бесцветные серобактерии окисляют сероводород:
H2S → S → – SO4.
Железобактерии переводят железо закисное в железо окисное:
Fe2+ → Fe3+.
Водородные бактерии окисляют молекулярный водород:
H2 → H2O.
Все эти процессы являются экзотермическими реакциями. Выделяемая энергия используется бактериями-хемосинтетиками для восстановления СО2 и синтеза органических соединений.
Хотя вклад хемосинтетиков в аккумуляцию энергии и синтез органических веществ на Земле невелик по сравнению с фотосинтезом, они имеют огромное экологическое значение, участвуя в круговороте веществ в биосфере.
2.5. Биологическое окисление
Анализируя отдельные этапы клеточного метаболизма, всегда необходимо помнить, что он представляет собой единый, интегральный, взаимосвязанный механизм (Бохински Р., 1987). Процессы анаболизма и катаболизма происходят в клетке одновременно и неразрывны друг с другом.
Анаэробный распад
Основным веществом, используемым клеткой для получения энергии, служит глюкоза. Анаэробный распад глюкозы – гликолиз, происходящий в цитоплазме клетки, является подготовительным этапом дыхания и основным этапом брожения. Поэтому мы можем сказать, что гликолиз – это один из важнейших процессов природы, свойственный всем организмам. Биохимия гликолиза хорошо изучена. Он включает 9 последовательных ферментативных реакций, конечным продуктом которых является пировиноградная кислота (ПВК):
С6H12O6 → 2C3H4O3.
Энергетическая эффективность гликолиза невысока и составляет 2 молекулы АТФ на одну молекулу глюкозы. Однако образованная ПВК – ключевое соединение в метаболизме клетки. Ее дальнейшее использование зависит от особенностей организма. У аэробных организмов ПВК включается в многоступенчатый процесс аэробного распада. В анаэробных условиях ПВК подвергается превращениям в различных видах брожения.