Глава 1
Основные принципы. Эволюция и естественный отбор. Обзор имеющихся на сегодня данных об эволюции высших приматов
Для того, чтобы говорить о чем-либо, следует вначале договориться об основных понятиях, установить правила игры, на которых затем мы будем строить свои рассуждения. Итак, объявляю эти основополагающие принципы.
1. Научный подход, дарвинизм. Такие теории происхождения человека, как «создание богом», «конструирование инопланетянами» и т. п. во внимание не принимаются.
2. Детерминизм. В широком смысле это означает, что у любого действия (события) есть причина, его породившая. В более узком, эволюционном, смысле это значит, что любое изменение внешнего и внутреннего строения живого существа, равно как и способов жизнедеятельности сообществ, имеет «железобетонную» причину, которая привела к этому изменению.
3. Естественный отбор. Любые изменения имеют адаптивный характер, то есть позволяют живым существам приспосабливаться к окружающей действительности. Закрепляются они естественным отбором, то есть тот, кто не обладает нужным признаком, вымирает. Следует обратить внимание, что, исходя из этого, абсолютно все эволюционные изменения происходят под «знаком смерти» (в том числе так называемый «половой отбор»). Другими словами, у любого изменения должна иметься не просто некая причина, но причина, угрожавшая самому существованию вида, – настолько, что все, кто не имел такого признака, был обречен на гибель (или на проигрыш во внутривидовой борьбе). Это же касается организации сообществ (стад, колоний и проч.). Выживают те, кто оптимально приспособлен к изменяющейся окружающей среде. Если исследователи не могут найти таких причин, то это вовсе не значит, что их не было. Скорее всего, просто плохо и не там искали.
Сюда же можно отнести такой принцип, как актуализированность адаптивных признаков. Для того, чтобы та или иная адаптация сохранялась длительное время, она должна быть востребована на протяжении всего этого временного отрезка. Не востребованный признак постепенно становится менее явным, а затем исчезает. В том случае, если не востребованный признак вдруг понадобился до своего исчезновения, он снова становится явным – происходит вторичная адаптация по данному признаку. Другой вариант – это появление каких-либо дальнейших изменений на основе данного признака.
4. Эволюционная достоверность. Если мы говорим о человеческих сообществах, то каждый период, каждая эпоха характеризуются не только определенными внешними признаками, но и особым инструментарием, с помощью которого человек познает и осваивает окружающий мир. Объясняя возникновение тех или иных социальных или индивидуальных черт, мы должны опираться на инструментарий, соответствующий эпохе, и не приписывать нашим далеким предкам образа мыслей и действий современных людей.
5. Независимость. Здесь в первую очередь имеется в виду то, что очень часто подобного рода исследования подгоняются под этическую и даже политическую коньюнктуру. Сознательно или бессознательно, исследователи, делая выводы, стараются придать им общий вид «политкорректности» и «благопристойности». Думаю, вряд ли наши далекие предки строили свою жизнь, сообразуясь с нынешними представлениями об общественной морали, у них и без того было достаточно проблем. Поэтому заранее приношу извинения особо чувствительным читателям за неприличное и асоциальное поведение наших далеких предков. Ей-богу, они никого не хотели обидеть.
6. Источники. Все используемые мной материалы общедоступны (и это принципиальная позиция). Это известные книги, переиздававшиеся множество раз (библиографию можно посмотреть в конце). Это также факты, хорошо знакомые абсолютно всем, начиная со школьного возраста. По большому счету, дело здесь не в фактах и источниках, а в том, как их расположить и в какую логическую последовательность выстроить. Если делать это беспристрастно и тщательно, последовательности возникают порой весьма неожиданные.
Прежде чем приступить к изложению наших соображений, следует вначале кратко упомянуть об общеизвестных в антропологической науке фактах. Приведем своего рода небольшой дайджест из общепринятых воззрений на происхождение нашего вида.
1
Итак, начало эволюционного пути человека можно отнести ко времени от 3,5 до 4 миллионов лет назад. В настоящее время признается, что эволюция гоминид была, скорее, не линейной, а кустообразной. Часто одновременно существовало по три, четыре, а, быть может, и более видов гоминид, в том числе на одной и той же территории.
Принято считать, что весь генезис гоминид (то есть прямоходящих) происходил на африканском континенте (ряд ученых это оспаривает). Именно Африка была тем плавильным котлом, в котором друг за другом возникали новые виды гоминид, и именно отсюда они затем расселялись по всему остальному земному шару, приспосабливаясь к конкретным условиям жизни в той или иной части мира и порождая внутривидовое и межвидовое разнообразие. Общепринято, что таких волн расселения было несколько. Относительно того, как именно и когда подобное расселение происходило, ученые ведут споры, и существует, по крайней мере, несколько гипотез на этот счет.
Как бы то ни было, новые виды, выходя из Африки, сталкивались на просторах Евразии с представителями более ранних форм «двуногих обезьян», вступали с ними в конкурентную борьбу, вытесняли из занимаемых ниш, либо скрещивались с ними, образуя гибридное потомство. Есть довольно значительное количество ученых, считающих, что человеческие расы образовались не только в результате адаптации под те или иные климатические условия и природные зоны, но как раз именно в результате подобного рода скрещивания (в последнее время генетические исследования подтвердили это предположение).
Предок человека довольно сильно отличался от представителя современного Homo sapiens. Собственно говоря, это даже еще не был «Homo» в полном смысле этого слова, а всего лишь его прапредок, которого антропологи назвали австралопитеком (в дословном переводе «южная обезьяна»). Именно к этой разновидности предков человека относилась знаменитая Люси – самка австралопитека, скелет которой, сохранившийся почти на 60 % (что весьма необычно для столь древних останков) раскопал в Юго-восточной Африке американский антрополог Д. Джохансон. Люси была предком как линии Homo, так и линии более поздних австралопитековых. Ей было присвоено родовое название «австралопитек афарский» (Australopithecus afarensis). Поскольку в числе останков сохранились тазовые кости и кости нижних конечностей, стало возможным утверждать, что уже на этой стадии существовало прямохождение. Соответственно, все потомки австралопитека афарского тоже были прямоходящими, или, как их стало принято называть, представителями «семейства гоминид». Примерно в период от 3 миллионов 200 тысяч до 3 миллионов лет назад линия ранних австралопитеков разделилась на линию собственно австралопитековых и линию Homo. Линию австралопитеков продолжил австралопитек африканский (Australopithecus africanus). Первым же в линии Homo стал так называемый «человек умелый» (Homo habilis).
2
Дальнейшее развитие линии австралопитеков пошло по пути укрупнения. Увеличивался их рост, становилась более массивной мускулатура, даже зубы стали более крупными. Большинство ученых считает, что это следствие пищевой специализации. У австралопитеков в рационе начинает преобладать растительная пища. Именно для пережевывания грубых растительных волокон им понадобились мощные жевательные мышцы, прикрепляемые к массивному костному гребню, а также большие коренные зубы. По своей пищевой специализации, а также по нише, занимаемой в окружающей среде, австралопитек стал походить на современных горилл. Так же, как у горилл, стал более заметен половой диморфизм. Самцы приобрели намного более крупные размеры по сравнению с самками, у них развились более крупные клыки (по всей вероятности, в качестве оружия в борьбе с другими самцами за обладание самками). Именно по этой причине такой австралопитек был назван антропологами «австралопитеком массивным» (Australopithecus robustus). Самой крупной разновидностью массивных австралопитеков был так называемый «австралопитек бойсовый» (Australopithecus boisei), его исследователи считают наиболее специализированым подвидом, той самой тупиковой ветвью, которая уже не могла приспособиться к изменившимся условиям окружающей среды. Ведь с одной стороны, специализация – вещь полезная, поскольку добавляет конкурентных преимуществ в краткосрочной перспективе, но с другой – вещь и очень опасная, поскольку узкая специализация предполагает максимальную адаптацию под конкретные условия окружающей среды. При резком изменении условий (например, климата, и, соответственно, сокращения пищевой базы) приспособиться специализированному виду будет очень сложно, а в большинстве случаев просто невозможно. Такой вид будет обречен на вымирание. Именно это и произошло с австралопитеками примерно 1,5–1,2 миллиона лет назад.
Впрочем, причины укрупнения австралопитеков многими оспариваются. Поводом к этому послужили последние исследования зубов австралопитеков – микроцарапин, возрастной изношенности, стертости эмали, а также ее изотопного состава. Анализ показывает, что диета массивных австралопитеков кардинально не отличалась от диеты австралопитеков грациальных. Основываясь на этом, ряд ученых предположил, что причиной укрупнения австралопитеков послужило вовсе не изменение пищевой специализации, а переход к иной социальной структуре по типу горилл, которые, как известно, придерживаются гаремного способа организации сообщества. Внутривидовая конкуренция между самцами при подобной организации сообщества многократно возрастает, что ведет к востребованности таких признаков, как масса тела, рост и наличие «оружия» в виде больших и острых клыков.
Во многом общественная жизнь австралопитеков остается до сих пор вопросом дискуссионным. Часть исследователей, как уже говорилось, считают, что социальная организация жизни австралопитеков походила на социальную организацию жизни горилл, то есть носила гаремный характер, но другая часть возражает, полагая, что социальная организация австралопитеков в гораздо большей степени напоминала быт стаи шимпанзе, где вполне могли уживаться сразу несколько самцов и самок.
Что касается орудий, то использование их австралопитеками не доказано. Нет ни одного более или менее убедительного свидетельства (находки), которая бы говорила нам об использовании австралопитеками (а главное – об изготовлении ими) орудий. Речь в первую очередь идет, конечно же, о каменных орудиях. Принято считать, что использование каменных орудий начинается на стадии «хабилиса». Тем не менее, ряд находок, сделанных в последнее время, все же склоняют исследователей к мнению, что австралопитеки могли время от времени использовать каменные орудия, имеющие природные сколы с острыми краями. Впрочем, в эпоху хабилисов изготовление каменных орудий сводилось к раскалыванию камня надвое путем бросания на твердую поверхность, чтобы получить острый край. Каменные орудия олдувайской эпохи – это всего лишь подходящие по размеру для руки гальки с острыми краями. Если обработка и присутствовала, то она была минимальной. Именно поэтому очень трудно судить о том, специально «изготавливались» эти орудия, или использовались уже готовые камни с острыми краями «природного» происхождения. Это касается орудий каменных. Но вполне очевидно, что орудия могли быть деревянными. О них, к сожалению, находки нам ничего не говорят. Да и не могут сказать, поскольку дерево не сохраняется на протяжении такого длительного времени. Нельзя с уверенностью также утверждать, что австралопитеки не пользовались костями с острыми сколами в качестве орудий. В пользу того, что использование как деревянных, так и костяных орудий вполне могло иметь место уже у австралопитеков, говорит хотя бы то, что даже обезьяны используют подобные орудия в процессе своей жизнедеятельности. Например, они пользуются палками для того, чтобы доставать фрукты, прибегают к помощи очищенных от коры веточек, чтобы вытащить термитов из термитника, пользуются также заостренными палками вроде дротиков, метая их в противника или в добычу. Некоторые исследователи отмечают, что обезьяны используют камни, чтобы раскалывать орехи и другую твердую пищу, а также кидаться ими в соперников. Но, как бы то ни было, речь идет лишь об использовании «подручных средств», а не о целенаправленном изготовлении орудий труда или охоты. Вероятнее всего, австралопитеки, несмотря на свою двуногость, орудий не изготовляли и широко их не использовали, по крайней мере так, как это делали представители линии Homo.
3
На стадии хабилисов начинается использование орудий труда. Постепенно начинается и их целенаправленное изготовление. Но пока это носит крайне примитивный характер.
Надо заметить, что ареалом расселения первых людей являлась достаточно ограниченная территория, которая включала в себя территорию нынешних Кении, Эфиопии и отчасти Танзании (примерно 2,4–2,0 млн. лет назад). Довольно долго «двуногие обезьяны» находились в пределах этого ареала. Существует общепринятое мнение, что ни австралопитеки, ни хабилисы не покидали африканский континент.
Примерно 3,4 миллиона лет назад началось изменение климата на африканском континенте, которое характеризовалось постепенным отступлением лесов и формированием относительно сухих пространств. Продолжалось оно почти полтора миллиона лет (до 2,0 млн. лет назад). Особо острый период изменения климата наступил 2,5 миллиона лет назад. К этому периоду относится эпоха переходного звена между австралопитеками и Homo, кости которого были найдены относительно недавно и который получил название Homo rudolfensis (существовал 2,4–1,9 млн. лет назад). Надо сказать, что в среде исследователей нет единства во мнении, кто именно был прямым предком хабилисов. Как бы то ни было, ясно лишь одно: в то время на африканском континенте имелось большое разнообразие гоминид. По саванне по-прежнему бродили австралопитеки – «массивные» и не очень, начал свое шествие и Homo habilis (1,9–1,6 млн. лет назад). Именно к тому времени относится вымирание большинства видов гоминид. Считается, что произошло это из-за резкого изменения климата и сокращения пищевой базы.
Многие ученые придерживается мнения, что популяция предков линии, которая привела к современному человеку, была весьма малочисленна – по разным оценкам, от 20 до 60 особей. Подобная теория получила название теории «бутылочного горлышка». Именно в масштабах этой популяции произошли серьезные эволюционные изменения, приведшие к возникновению нового вида. Изменения эти не были одномоментными и продолжались длительное время (после прохождения «бутылочного горлышка») – вплоть до появления Homo erectus, следующей ступени в развитии предков человека. «Бутылочное горлышко» на протяжении эволюции предков человека возникало не один раз. Эволюционный скачок, то есть серьезные эволюционные изменения, происходящие в сжатый временной отрезок, практически всегда происходит в небольших популяциях, которые в итоге оказываются наиболее приспособленными и с течением времени распространяют вновь приобретенные признаки на всех своих уже гораздо более многочисленных потомков.
Какие же изменения претерпел человек на стадии хабилиса и следовавшего за ним эректуса?
1. Радикальная перемена пищевого рациона и пищевой стратегии (переход на животную пищу).
2. Совершенствование локомоции (движения) на основе бипедии (прямохождения).
3. Начало систематической обработки камня.
Говоря о переходе на животную пищу, ученые сходятся во мнении, что гоминиды поначалу были падальщиками, поскольку вряд ли сами были способны охотиться на крупных животных. Однако говоря, что гоминиды были падальщиками, ученые вовсе не имеют в виду, что они питались полуразложившимися трупами животных. Речь идет о доедании добычи за крупными хищниками. Был даже введен специальный термин, обозначающий подобного рода пищевую стратегию – «скэвинджинг». При этом выделяются активный скэвинджинг и пассивный. При пассивном пищевая стратегия состоит в том, чтобы дождаться окончания трапезы крупных хищников и заполучить остатки. Надо заметить, что в тот период стада крупных копытных в Африке были довольно многочисленны, после хищников оставалось много мяса, так что подобная стратегия вполне могла быть успешной. Кроме того, гоминиды при помощи каменных орудий вполне могли дробить кости животных для того, чтобы добыть костный мозг – весьма ценный в энергетическом смысле продукт. Единственный, кто мог в этом составить им конкуренцию, – гиены. Однако гиены были распространены не на всей территории африканского континента (в основном к югу), в отдельных районах их присутствие было весьма ограниченным, так что пищевая ниша была малоконкурентна. Кроме того, гиены и прочие хищники в основном активны ночью и в сумерках, а Homo, по мнению ученых, занял нишу «полуденного падальщика». От пассивного скэвинджинга гоминиды постепенно переходили к скэвинджингу активному. Имеется в виду та стратегия, когда хищников уже начинают отгонять от добычи.
4
Как уже отмечалось, использование орудий (и в этом также сходятся почти все исследователи) может оказаться не только прерогативой представителей рода Homo. В последнее время сделан ряд находок, которые вполне можно охарактеризовать как орудия, но которые при этом имеют слишком большой возраст (напр., раскопки в Када Гона, Эфиопия, возраст 2,6–2,9 млн. лет) и не могут являться «делом рук» Homo, поскольку его тогда просто не существовало. Из-за датировки многие склонны приписывать использование этих орудий австралопитекам.
Если использование орудий вполне может оказаться свойственным не только человеку, то осознанное, систематическое изготовление орудий и их использование свойственно исключительно представителям рода Homo.
Homo erectus (человек прямоходящий) – следующая ступень в становлении рода человеческого.
К переходной форме от хомо хабилис к хомо эректус принято относить так называемых «людей из Дманиси», останки которых были обнаружены в Грузии в конце XX века. Датируются они 1,77 млн. лет назад. Это наиболее древние свидетельства миграции предков людей за территорию Африки. Наряду с прогрессивными признаками посткраниального скелета (то есть всего остального скелета, кроме черепа), «люди из Дманиси» имеют и архаичные черты, сближающие их с хабилисами. В первую очередь это малый объем мозга, практически не превышающий объем мозга хабилисов. Вместе с костями «людей из Дманиси» было обнаружено довольно много костей всевозможного крупного и мелкого зверья. На некоторых остались царапины, оставленные каменными орудиями. Одна кость большого травоядного животного была разгрызена крупным хищником уже после того, как люди соскребли с нее мясо. Это, конечно же, не является свидетельством того, что древние люди умели охотиться, но вполне четко указывает, что люди могли получать доступ к тушам раньше своих конкурентов – медведей, гиен, леопардов и саблезубых тигров. Орудия, найденные в Дманиси, самые примитивные – олдувайские.
После появления в Грузии древние люди начали расселяться по югу Азии. На крайний юго-восток континента эректусы проникли, судя по всему, около полутора миллионов лет назад. Спустя примерно еще 200–300 тыс. лет, они появились на северо-востоке Китая. Многие ученые считают, что эти две популяции эректусов (китайская и юго-восточная) жили изолированно друг от друга, поскольку их разделяли густые субтропические леса. Леса эти были населены орангутанами, гиббонами, вымершими огромными обезьянами гигантопитеками и гигантскими пандами. Леса эти были непригодны для жизни эректусов, которые предпочитали открытые пространства. Северо-китайская популяция приспособилась к жизни в сухом и относительно прохладном климате, ныне она известна под названием синантропов (вымерли 400–350 тыс. лет назад). Южная популяция приспособилась к жизни в экваториальной зоне и просуществовала значительно дольше: последние представители эректусов, вполне вероятно, жили на острове Ява всего 50 тыс. лет назад.
В целом период существования Homo erectus довольно продолжительный, поэтому его принято разбивать на несколько частей, в соответствии с теми изменениями, которые происходили как во внешнем облике, так и в поведении представителей данного вида.
Наиболее ранней, архаичной формой Homo erectus является так называемый Homo ergaster (человек работающий), время существования 1,8–1,5 млн. лет назад.
Более поздняя и более продвинутая форма Homo erectus – Homo antecessor (человек предшествующий), время существования от 1,2 млн. до 800 тыс. лет назад.
Еще более поздним подвидом Homo erectus является Homo heidelbergensis (Гейдельбергский человек), время существования 800–345 тыс. лет назад (ряд ученых, впрочем, относят его к «преднеандертальцам», таким образом выводя за пределы вида Homo erectus). Относительно того, кто является непосредственным предком человека разумного, ученые спорят. Одни считают, что таковым является гейдельбергский человек, другие приписывают эту честь виду Homo antecessor, третьи считают, что этого представителя ископаемых гоминид еще только предстоит найти.
Другими словами, в этот период гоминиды снова очутились на распутье, в результате чего возникло две разновидности людей: неандерталец и кроманьонец (то есть человек современного типа). Однако прежде следует подвести итоги того пути, который гоминиды проделали на стадии человека прямоходящего. Что же произошло знаменательного на этом этапе?
1. Еще большее овладение техникой прямохождения, развитие выносливости, способности передвигаться на большие расстояния, бегать. Развитие адаптационных функций организма, сопутствующих этому: терморегуляции, амортизационного изгиба позвоночника и арки стопы и т. п.
2. Окончательная утрата волосяного покрова.
3. Совершенствование орудий труда. Переход от примитивной Олдувайской культуры обработки камня (характерной для хабилисов) к более продвинутой Ашельской культуре. Начало изготовления не только орудий, но и оружия, годного для того, чтобы самостоятельно охотиться. Анализ каменных орудий человека прямоходящего показал, что уже ранняя его форма (Homo ergaster) использовала каменные орудия для того, чтобы обрабатывать дерево.
4. Освоение огня.
5. На стадии Homo erectus происходит увеличение головного мозга почти в два раза (по сравнению хабилисом) – приблизительно от 650 до 1000 сант. куб.
6. Происходит активная экспансия человека прямоходящего по земному шару: вначале по африканскому континенту, а затем и за его пределами. Эректус заселяет восточную, западную и северную часть Африки, в наиболее узких местах Красного моря (Суэцкий перешеек и Баб-эль-Мандебский пролив) перебирается в Азию и, двигаясь вдоль побережья, достигает территорий современной Северной Индии и Китая. Следуя по череде многочисленных островов, оказывается в Индонезии (Ява).
Расселившись по всему свету, эректусы, естественно, адаптировались к тем условиям, в которые попадали. Так в 2004 году было опубликовано описание неизвестного ранее вида ископаемых людей Homo foresiensis, живших на острове Флоренс (Индонезия) еще около 20 тыс. лет назад. Примечательным в этом виде было то, что рост найденных индивидуумов не превышал метра. Объем черепной коробки тоже был очень мал, что-то около 400 куб. сант. В посткраниальном скелете, в частности, в строении плечевого сустава, присутствовало множество архаичных черт. Новый вид получил обиходное название «хоббитов» и вызвал в среде ученых ожесточенные споры. В конце концов было выработано общее мнение, что «хоббиты» являются потомками эректусов, попавших на остров примерно 800 тыс. лет назад, или ранее. За сотни тысяч лет спокойной жизни среди карликовых слонов (стегодонов), варанов и крыс островитяне измельчали, а технологическое развитие каменных орудий у них практически полностью остановилось. Подобное измельчение довольно часто происходит с оказавшимися в островной изоляции животными. Подобное произошло, например, с ископаемыми карликовыми гиппопотамами, жившими на острове Мадагаскар еще сравнительно недавно (где-то тысячу лет назад). Подобно предкам «хоббитов», они, попав на остров, стали мельчать, при этом их мозг уменьшался гораздо быстрее, чем это можно было ожидать.
Появление на острове людей современного типа (рослых верхнепалеолитических охотников с высокоразвитой каменной индустрией) обрекло проживший множество лет в изоляции народ на вымирание. Впрочем, среди современных островитян бытуют легенды о маленьких лесных людях, так что последние «хоббиты», вполне вероятно, вымерли всего несколько столетий назад.
Надо заметить, что учеными высказывалось и иное мнение относительно происхождения «хоббитов». Ряд ученых считает, что «хоббиты» произошли от более ранних гоминид – ранних эректусов, близких к людям из Дманиси, или даже хабилисов. По их мнению, это гораздо лучше согласуется с маленьким размером мозга особей данного вида.
Как бы то ни было, пример «хоббитов» со всей очевидностью свидетельствует о том, что эректусы расселились практически по всему миру, проникнув такие места, где до них не бывал ни один из видов гоминид. По существу, эректусы совершили масштабную экспансию в мировом масштабе. Однако, говоря об экспансии, следует учитывать, что она не носила одномоментный характер. Это не была одна группа, которая вышла за пределы африканского континента и совершила длинный, периодом в несколько тысяч лет марш-бросок, попутно размножаясь и заселяя встречающиеся по дороге пространства. Экспансия проходила волнами, причем таких волн было множество. С африканского континента по протоптанной дорожке приходили все новые представители человека прямоходящего. Они встречались с теми, кто уже жил в этих местах, смешивались с ними, возможно, воевали, шли дальше. При этом эволюция не стояла на месте. Пришельцы вполне могли уже отличаться от тех, кто жил в пределах новых территорий. В процессе конкурентной борьбы они могли вытеснять тех, кто пришел сюда раньше. Вероятно, что-то подобное произошло с Гейдельбергским человеком (Homo heidelbergensis) и Человеком предшествующим (Homo antecessor) – как мы помним, поздними формами эректуса (т. е. человека прямоходящего), последовательно появлявшимися на территории Евразии.
5
Как шла эволюция от эректусов к сапиенсам, известно довольно хорошо. По крайней мере, лучше, чем многие другие более древние узловые моменты эволюции. Коротко напомним то, что к настоящему времени ученым удалось установить.
После эректусов хронологически идет так называемый «денисовский человек». Известен он по крайне фрагментарному материалу, обнаруженному в Денисовой пещере, расположенной в России, в Алтайском крае (2008 г.). Предположительно время существования денисовца относится к 800-40 тыс. лет назад. Из-за скудости находок (известно всего пять фрагментов: зубы и фаланги пальцев) восстановление облика денисовского человека пока не представляется возможным. Тем не менее, этих находок было достаточно для того, чтобы произвести генетические исследования. Исследования генома показали, что денисовцы были темнокожие, темноглазые, темноволосые. Денисовские люди стали вторым после неандертальцев видом вымерших гоминид, для которого стал известен полный митохондриальный и почти полный ядерный геномы (2012 г.). Фактически данный вид приматов был выделен исключительно на основании генетических исследований. После подробного изученя ядерного генома, а также проведения радиоуглеродного исследования, выяснилось, что разные костные остатки датируются периодами от 170 до 50 тыс. лет назад, а кроме того, обнаружилось, что геном «денисовца» похож на геном гейдельбергского человека (Homo heidelbergensis). По данным генетического исследования расхождение ветвей гейдельбергского человека и денисовца произошло около 700 тыс. лет назад, а с неандертальцем денисовцы разделились около 500 тыс. лет назад. Их общий предок (неандертальца и денисовца соответственно) с представителями ветви, ведущей к нашему собственному виду (Homo sapiens) разошелся примерно 700–765 тыс. лет назад.
Генетические исследования также показали, что для денисовцев была характерна особая вариация белка EPAS1, участвующая в кислородозависимой регуляции некоторых генов. Такая же вариация встречается у современных тибетцев (у других популяций людей ее нет), что, вполне вероятно, им позволило приспособиться к жизни на большой высоте. Скорее всего, эта генетическая особенность им досталась в наследство от денисовцев.
Следы гибридизации денисовского человека с Homo sapiens (современным человеком), как оказалось, присутствуют в геноме в достаточно большом объеме. Так в геноме меланезийцев обнаружено около 5 % общих генов с прочитанным ядерным геномом денисовского человека. В то же время общих генов у современного человека и неандертальца от 1 до 4 % в различных популяциях (за исключением коренных жителей Африки, у которых неандертальские гены отсутствуют). Неандертальское «наследство» наиболее выражено в генах жителей Европы и Пакистана, денисовское – у обитателей южно-тихоокеанского региона (т. е. меланезийцев, представителей австралоидной расы, среди которых у нас наиболее известны папуасы), что косвенно указывает на изначальное территориальное распределение этих видов. Рядом ученых предполагается, что появился денисовский человек из Африки, причем это была волна миграции, отдельная от волн эректусов, неандертальцев и сапиенсов. Согласно ДНК-исследованиями 2013 года, установлено, что неандертальцы скрещивались с кроманьонцами и денисовцами, кроманьонцы – с денисовцами, а кроме того, денисовцы скрещивались с каким-то четвертым, ранее неизвестным видом людей (остатков которых пока не найдено, но «следы» генов которых присутствуют в геноме денисовца). Некоторые ученые считают потомком денисовцев так называемого «человека из оленьей пещеры», обнаруженного на юге Китая в 1979 году (черепа из скальной породы были извлечены лишь в 2009 году). Возраст останков определяют от 11,5 до 14,3 тыс. лет. Это ныне вымершая популяция людей. У обладателей извлеченных из пещеры черепов была круглая черепная коробка, низкие надбровные дуги, небольшое плоское лицо с широким носом, сильно выдающаяся вперед нижняя челюсть. Останки отличает сочетание признаков, свойственных современному человеку, и черт заметно более архаичных. Это самые поздние известные науке останки группы людей, которая не вписывается в рамки изменчивости Homo sapiens и, возможно, принадлежит другому виду рода Homo. Некоторые ученые склоняются к тому, что это изолированная популяция современных людей, приобретшая благодаря длительному автономному существованию специфические черты (из-за так называемого дрейфа генов), другие, как уже отмечалось, полагают, что это популяция, возникшая при скрещивании денисовцев с предками австронезийцев.
Как свидетельствуют находки в Денисовой пещере, у денисовцев была довольно развитая материальная культура. Это миниатюрные каменные иглы из костей птиц с просверленным ушком, бусины из скорлупы страусиного яйца, ожерелья из зубов животных, подвески из ракушек, украшения из поделочного камня. Был найден даже женский браслет, сделанный из довольно хрупкого камня, ближайшие выходы пород которого находятся за двести километров от пещеры. При этом для обработки камня применялись приемы, совершенно нетипичные для ранней стадии верхнего палеолита (станковое сверление, внутренняя расточка, шлифование и полировка). Набор технических приемов, применявшихся денисовцами более 45 тыс. лет назад, у людей современного вида типичен для гораздо более поздней эпохи – бронзового века.
Если денисовцы на генеалогическом древе занимают положение отдельной ветви, то другое ответвление, ведущее к неандертальцам, проходит, вероятнее всего, через стадию гейдельбергского человека (см. рис 1).
Гейдельбергский человек – это исключительно европейский вариант Homo. Значительное число ученых считают его разновидностью человека прямоходящего (как китайского синантропа и индонезийского питекантропа). Обитал он в Европе с 800 до 345 тыс. лет назад (некоторые ученые датируют исчезновение гейдельбергского человека более поздним периодом, доводя его существование до 130 тыс. лет назад). Вероятнее всего, гейдельбергский человек является потомком европейского «человека-предшественника» (Homo antecessor, поздняя форма эректуса) и в то же время – непосредственным предшественником неандертальца. Как уже говорилось выше, после изучения ядерной ДНК гейдельбергского человека (материал был взят от трех образцов, найденных в одной из испанских пещер) выяснилось, что гейдельбергцы находились на линии, ведущей к неандертальцам. Эволюционное расхождение ветви денисовцев с ветвью общей для неандертальцев и гейдельбергцев произошло, по оценкам ученых, около 500 тыс. лет назад (точнее – ранее 430 тыс. лет назад). Общий же предок Homo heidelbergensis и Homo sapiens жил 700–765 тыс. лет назад.
Находки останков гейдельбергского человека были сделаны в Германии, Италии, Франции, Греции, Испании, Великобритании.
В целом морфология гейдельбергского человека мало отличалась от морфологии эректуса. Впрочем, рост его не превышал 1,5 метра. От предшественников он отличался некоторыми прогрессивными признаками (порой свойственными конкретно неандертальцам), в частности большим мозгом и округлым затылком, а от последующих, более продвинутых форм, – архаично-низким черепом с толстыми стенками и очень массивными челюстями с большими зубами.
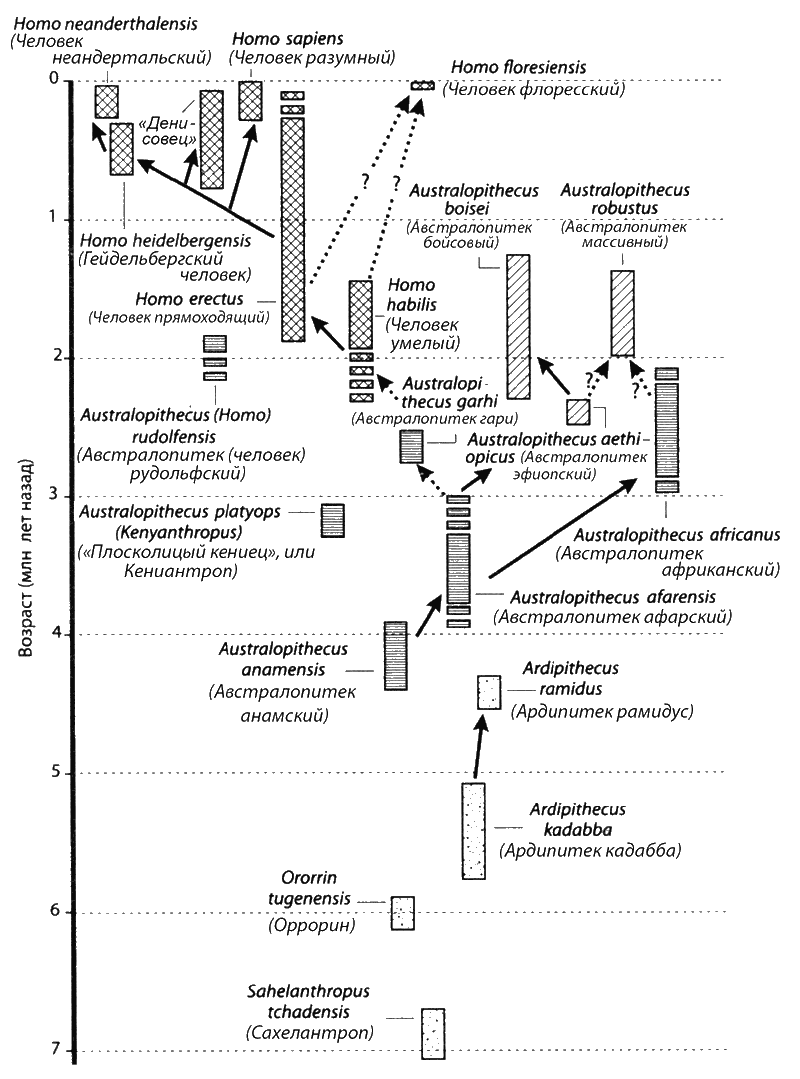
рисунок 1
Изменения коснулись мозга гейдельбергского человека. Выросла высота лобной доли, развилась теменная область (ее размеры находятся вне пределов изменчивости эректуса). Височная доля имела уже вполне современные пропорции, хотя и меньшие, чем у современного человека, размеры. Затылочная же доля и мозжечок не изменились со времен существования эректуса. В целом форма мозга гейдельбергского человека свидетельствует о резком прогрессе в области контроля за движениями, в том числе способностей к прогнозированию и планированию своих будущих действий. Необходимо также отметить бурное развитие области, обеспечивающей согласование речи и движений рук, а также рельефное выступание зоны Брока, свидетельствующее о начале, как считает часть ученых, использования речи.
Культура гейдельбергского человека характеризуется как шелльская (аббевильская), основными орудиями которой являются ручные рубила, отщепы, чопперы и проч. В их изготовлении заметен серьезный прогресс, выразившийся, в частности, в двусторонней симметричности ручных рубил. Ряд ученых полагает, что гейдельбергский человек мог строить примитивные хижины и пользоваться огнем (впрочем, другими учеными это опровергается, поскольку ископаемые свидетельства этих фактов они относят к эпохе неандертальцев). В 90-е года XX века в Германии (в шахте по добыче угля в Шёнингене) были найдены копья из дерева (8 штук). Копья датируются около 300 тыс. лет назад и являются самыми древними полностью сохранившимися образцами оружия для охоты. Они являются также важнейшим доказательством «осознанной охоты» гейдельбергского человека. Шёнингенские копья позволяют предположить, что гейдельбергские люди охотились с помощью деревянных копий даже на крупных животных, однако мясо ели сырым, поскольку следов огня на стоянках не обнаружено. Для изготовления копий использовались сосна и ель, их длина находилась в пределах между 1,8 и 2,5 м. Копья тщательно обработаны и свидетельствуют о высоком технологическом мастерстве и ремесленной традиции. Центр тяжести копий находится в передней трети, как и у современных копий для метания, используемых при спортивных состязаниях. Были проведены специальные эксперименты, во время которых спортсменам удалось бросить точные копии этих орудий на расстояние в 70 метров. Вместе с копьями было найдено более чем 10 тыс. останков костей животных, 90 % из которых относятся к диким лошадям, остальные 10 – это в основном благородный олень и бизон. Останки лошадей носят следы разрезов, сделанных каменными орудиями, и лишь немногие – следы от зубов хищников. Кроме того, были найдены: обуглившаяся палка (интерпретируемая некоторыми учеными как «вертел»), а также с обоих концов заостренное деревянное орудие, назначение которого до конца не прояснено. В любом случае данные орудия интерпретируются как имеющие отношение к охоте и разделке мяса.
На севере Испании с 1994 года ведутся раскопки в пещере Атапуэрка, где обнаружены останки 28 особей, относящихся к виду «гейдельбергский человек» (до этого по всей Европе обнаружены останки всего 26 особей, относящихся к данному виду). Останки были найдены на дне 14-метровой ямы и датируются возрастом около 350 тыс. лет. Две трети погибших – это особи в возрасте от 11 до 20 лет, среди них нет ни одного детского скелета (в ту эпоху люди жили не долго), зато чуть более половины – женщины. Все это вкупе дало ученым повод говорить о том, что речь идет о нападении одного племени на другое и последующей расправе над побежденными. Кроме того, в ходе раскопок были обнаружены свидетельства каннибализма гейдельбергцев, поскольку мясо с костей, принадлежащих десяти представителям доисторического человека, было срезано специальными резаками. Ученые допускают, что это было сделано не только в пищевых, но и в ритуальных целях.
6
Прямым потомком гейдельбергского человека является неандерталец. Пожалуй, в настоящее время это самый изученный вид ископаемых гоминид. Первые люди с чертами протонеандертальца появились в Европе около 600 тыс. лет назад. А последние из неандертальцев населяли континент всего 25 тыс. лет назад. Время же существования классических неандертальцев укладывается в промежуток от 350 до 120 тыс. лет назад. Первый представитель этого вида был найден в ущелье Неандерталь близ Дюссельдорфа (западная Германия) в 1856 году. В последующие 150 лет останки неандертальцев находили в большом количестве по всей Европе: в Германии, Франции, Италии, Греции, в Крыму. Неандерталец был также обнаружен на Кавказе, в Средней Азии, на Алтае, Ближнем и Среднем Востоке. Соответственно, ареал его распространения не ограничивался только Европой, а частью захватывал территорию Азии. Довольно значительное время неандертальцы сосуществовали с кроманьонцами на одних территориях, а в некоторых случаях поочередно занимали одни и те же стоянки. Например, на территории Израиля обнаружены пещеры, которые несколько раз переходили «из рук в руки»: до 130 тыс. лет назад там жили неандертальцы, со 130 до 80 тыс. лет – кроманьонцы, с 65 до 47 тыс. лет назад – снова неандертальцы, а затем опять люди современного типа.
В целом неандертальцы внешне похожи на людей современного типа, однако есть ряд существенных особенностей, свойственных только им. Средний рост неандертальца составлял около 165 см., половой диморфизм у них не был выражен ярко и находился примерно в рамках тех же пропорций, что и у современного человека (самка где-то на 10 см ниже самца). Объемы эндокранов говорят о том, что размером мозга неандертальцы даже превосходили современного человека (1400–1740 см куб. против 1350–1400 см куб.), при этом форма мозга неандертальца существенно отличалась от мозга кроманьонца. Если у современного человека объем мозга возрастал по сравнению с предшественниками в основном за счет коры больших полушарий, лобной и теменных областей, то у неандертальца – за счет затылочной части. Черепная коробка у неандертальца поэтому длинная и широкая и по сравнению с нашей низкая и уплощенная. Вообще кости черепа (как и все остальные) у неандертальца более массивные, чем у современного человека. Лоб – покатый, имеются ярко выраженные, мощные надбровные дуги, которые, сливаясь над переносицей, образуют сплошной валик, окружающий глазницы сверху и с боков. Характерной неандертальской особенностью является ярко выраженный затылочный валик, а также (особенно хорошо заметный в профиль) шиньонообразный выступ в верхней части затылка. Отверстие в нижней части мозговой коробки, соединяющее полость черепа с позвоночным каналом, у неандертальцев имеет не округлую форму, как у нас, а удлиненно-овальную. Неандертальцу также был свойственен так называемый среднелицевой прогнатизм (т. е. «выдвинутость вперед» средней части лица). Нос у них был большой (носовое отверстие в черепе было также намного крупнее и шире нашего), верхнечелюстные пазухи (по бокам от ноздрей) сильно раздуты, а челюсти несколько выдавались вперед. Подбородочный выступ у неандертальца отсутствовал, отчего его подбородок был как бы «срезан».
Посткраниальный скелет также имел ряд специфических особенностей. Шея неандертальца была короткая и наклоненная вперед, грудная клетка имела колоколовидную, т. е. расширяющуюся книзу форму, а не бочкообразную, как у современного человека. Ребра он имел более толстые, руки и ноги – укороченные по сравнению с туловищем. Кости неандертальских конечностей (как верхних, так и нижних) выделяются крупными суставными головками, большей толщиной стенок и более значительной рельефностью поверхности (из-за лучшей выраженности участков крепления мускулов). Бедренная и лучевая кости неандертальца более изогнуты, чем у современных людей, плечи – шире, таз был также шире и длиннее, чем у сапиенсов. Мышечная масса неандертальца была на 30–40 % больше, чем у кроманьонца.
В целом особенности строения скелета неандертальца объясняются адаптацией данного вида к суровым северным условиям, а также большим физическим нагрузкам, которые в течение жизни переносили их обладатели. Не исключено, что некоторые из этих особенностей не были предопределены на генетическом уровне, а образовывались (или усугублялись) в течение жизни вследствие час тых и долгих переходов, транспортировки тяжестей без вспомогательных средств, выслеживания и преследования охотничьей добычи, столкновений с крупными опасными животными и т. д.
Цвет кожи неандертальца, по всей видимости, был таким же, как у современных жителей Европы. Дело в том, что в высоких широтах иметь темную кожу невыгодно: солнца и так мало, а она его «отталкивает» и, соответственно, мешает выработке необходимого организму витамина «Д». Кроме того, о светлокожести неандертальцев свидетельствует их ДНК. Дело в том, что ген меланокортина (mc1r), выявленный в ДНК неандертальцев, живших в Испании примерно 40–50 тыс. лет назад, показал, что если и не сами обладатели этих генов, то многие их соплеменники были светлокожими и рыжеволосыми.
Средняя продолжительность жизни неандертальцев составляла около 23 лет (половая зрелость у них наступала уже к 8-10 годам). Это не значит, что среди них не было пожилых, подобная цифра получилась вследствие крайне высокой детской смертности (более 50 %). Рожали неандерталки (это также установлено исследованиями) в среднем раз в 3–4 года. Многие останки неандертальцев носят на себе следы гипоплазии (нарушение в структуре зубной эмали), по всей видимости, являющейся следствием недоедания и болезней в период ее формирования. Наконец, на многих неандертальских костяках есть следы переломов и других многочисленных травм.
Идентичность гена FOXP2 (связанного с речью) у современного человека и неандертальца, а также строение голосового аппарата и мозга неандертальцев позволяют сделать вывод о том, что они могли обладать чем-то вроде речи. Некоторые ученые полагают, что такой речью мог быть «язык из согласных с малым количеством гласных, что встречается и в человеческих языках».
Диета неандертальцев была мясной (об этом свидетельствуют специальные данные исследований костной ткани). Однако, как показало изучение их фекалий, в рацион также входили ягоды, орехи и клубни. При том, что холодный климат требовал от неандертальцев больших затрат энергии на поддержание температуры тела, короткие конечности при передвижении также сжигали больше калорий, чем, скажем, конечности кроманьонца (примерно на 32 %). Соответственно, ежедневная потребность в пище неандертальца, по мнению некоторых ученых, была выше, чем у кроманьонца (примерно на 100–350 килокалорий), если бы он, конечно, обитал в тех же широтах.
Как уже говорилось выше, неандерталец и денисовец – это на настоящий момент те из ископаемых гоминид, которые подверглись генетическим исследованиям. Точнее, из их останков было возможно выделить генетический материал, пригодный для идентефикации. Исследователи извлекли ископаемую ДНК из кости бедра неандертальского скелета 38 тысячелетней давности из пещеры Виндия в Хорватии, а также из других костей, найденных в Испании (Эль-Сидрон), России (Мезмайская пещера) и Германии (Фельдхофер). Древнейший неандерталец, из которого удалось извлечь генетическую информацию, это человек из Альтамуры (Италия), живший 130–172 тыс. лет назад. Геном неандертальца по размеру близок к геному современного человека. Ученым удалось восстановить 63 % неандертальского генома (2 миллиарда пар нуклеотидов, если исключить повторяющиеся фрагменты), а оставшиеся 37 % его длины (1,2 миллиарда пар нуклеотидов) еще не выявлены. Предварительные результаты показывают, что ДНК современного человека и неандертальца идентичны приблизительно на 99,5 %.
Прочтение генома неандертальца состояло из двух этапов. Вначале была прочитана митохондриальная ДНК. Из курса средней школы всем хорошо известно, что митохондрии – это симбиотические органеллы, находящиеся в цитоплазме клетки, получающие от нее все необходимые питательные вещества, а взамен поставляющие ей энергию. Они имеют свою собственную хромосому, поскольку изначально были отдельными одноклеточными организмами, а затем в результате эволюции выбрали местом своего обитания «внутренности» клетки. Митохондриальная ДНК намного меньше ДНК, содержащегося в ядре клетки, поэтому ее гораздо легче прочесть. Кроме того, при половом размножении слияние отцовской и материнской ДНК происходит лишь в ядре, митохондриальная ДНК остается постоянно неизменной и наследуется по материнской линии. К слову, именно это обстоятельство позволило ученым проследить «путь» митохондриальных ДНК на протяжении последних 150–200 тыс. лет и на основе подсчета и анализа происходящих в них мутаций выяснить, от какого количества «праматерей» произошло все современное человечество. Вывод был следующим. Все современное человечество произошло от относительно малочисленного сообщества, которое обитало в юго-восточной Африке и насчитывало всего 7 самок. Время жизни «митохондриальной Евы», как стали называть всеобщую «праматерь», было определено в 180–200 тыс. лет назад, впрочем, более поздние исследования скорректировали эту цифру и определили в 140–120 тыс. лет.
Примерно такие же манипуляции чуть позже были произведены с Y-хромосомой, которая, как известно, есть только у мужчин и, соответственно, наследуется только по мужской линии – от отца к сыну. Однако Y-хромосома находится в ядре клетки, она намного длиннее митохондриальной ДНК (примерно на 60 миллионов пар оснований), мутации в ней накапливаются гораздо медленнее, что снижает точность оценки. Тем не менее, после долгих подсчетов разные группы ученых пришли к той же самой цифре: «Y-хромосомный Адам» жил в промежутке от 140 до 120 тысяч лет назад. Но, как мы понимаем, речь в данном случае идет о «сапиенсах».
При прочтении генома неандертальца вначале тоже была прочитана митохондриальная ДНК, а затем – ядерная. Анализ полученных данных, в частности, показал, что генетически пути неандертальцев и современного человека разошлись ранее 500 тыс. лет назад, то есть еще до распространения ныне существующих рас человека.
Ученым удалось узнать очень много о генеалогических связях неандертальца, о его морфологии, им даже удалось обнаружить ген, препятствующий усвоению молока (лактозы) в зрелом возрасте. Также в ходе исследований выяснилось, что неандертальцам были незнакомы многие наследственные заболевания современных людей – аутизм, болезнь Альцгеймера, синдром Дауна, шизофрения.
В настоящее время точно установлено, что неандерталец не был предком человека, это подтвердило исследование его митохондриальной ДНК и сравнение с митохондриальной ДНК современного человека. Как уже говорилось, примерно 700–760 тыс. лет назад от общей ветви, идущей предположительно от эректуса, отпочковалась ветвь, ведущая к современному человеку. Около 700 тыс. лет назад другая линия пошла к денисовцу. Третья ветвь (около 500 тыс. лет назад) через гейдельбергского человека вела к неандертальцу. Предки нашего вида пришли из Африки всего около 40 тыс. лет назад, в то время как в Европе на протяжении десятков тысяч лет безраздельно царствовали неандертальцы. В отдельных регионах неандертальцы и человек разумный сосуществовали многие тысячи лет. Некоторые антропологи считают, что два вида людей преимущественно враждовали. Так на стоянках и неандертальцев, и кроманьонцев были обнаружены обглоданные кости другого вида. Также имеются сведения, что кроманьонцы делали ожерелья из зубов неандертальцев. В погребальном комплексе кроманьонцев Сунгирь (на окраине города Владимира в 200 км от Москвы) была найдена большая берцовая кость неандертальца с отсеченными суставами, полость которой содержала порошок охры (соответственно, кость использовалась как своеобразная шкатулка). Другие ученые предполагают, что между двумя видами был более мирный характер взаимоотношений. Как уже говорилось, имеются генетические и антропологические подтверждения смешения между неандертальцами и современными людьми. Генами неандертальцев в разной степени (от 1 до 4 %) обладает практически все население Земли, исключая коренных жителей Африки (что недвусмысленно указывает: метисизация происходила за пределами данного континента). Так в 2002 в румынской пещере Пештера-ку-Оасе была найдена кость человека современного вида, жившего около 40 тыс. лет назад и принадлежавшего к первой волне кроманьонцев, появившихся в Европе. Согласно опубликованным в 2015 году результатам ДНК-исследований, от 6 % до 9 % его генома было унаследовано от неандертальцев и от неандертальского предка его отделяло всего 4–6 поколений. В Рипаро ди Меззена (север Италии) найдены, предположительно, останки гибрида неандертальца и Homo sapiens, жившего ок. 30–40 тысяч лет назад. В Староселье (Крым) было обнаружено погребение ребенка вида Homo sapiens с отдельными неандерталоидными признаками.
Ряд ученых предполагает, что неандертальцы жили не только в пещерах, но еще умели возводить примитивные жилища. Раскопками установлено, что жилище неандертальцев делилось на три «зоны». В первой из них располагалась «кухня», о чем свидетельствуют найденные в этой зоне останки животных и следы охры, которая применялась для выделки шкур, в качестве клея и как антисептическое средство. Во второй зоне, которая была самой просторной, размещался очаг и здесь было спальное место. В третьей зоне была «мастерская», где изготавливались каменные инструменты, здесь было больше всего солнечного света и минимальный риск, что кто-то помешает работе.
Как явствует из описания, неандертальцы уже вполне «приручили» огонь (хотя мясо ели в основном в сыром виде). Возможно, огонь использовался ими для других целей: например, для обогрева жилища, для избавления от паразитов при помощи дыма и др. Относительно социальной структуры общества у неандертальцев можно сказать, что они жили сообществами по 10–20 особей. Некоторые полагают, что особи мужского пола оставались в племени, в то время как особи женского пола переходили в племя мужа (об этом говорит ряд косвенных свидетельств, полученных при генетическом исследовании родственных связей живших в одной пещере неандертальцев).
Никаких свидетельств существования культуры и искусства у неандертальцев нет. То, что поначалу было принято за костяную флейту с 4-мя отверстиями, в итоге оказалось просто следами зубов хищника (гиены). В ряде пещер в Бельгии, Франции, Израиле, Узбекистане, Ираке были найдены останки неандертальцев, имеющие на себе признаки погребений. Именно таким образом зачастую истолковывали их исследователи. Здесь следует заметить, что большинство таких находок делались в то время, когда неандерталец считался непосредственным предшественником человека. Отсюда становится понятным стремление некоторых ученых выдать желаемое за действительное, увидев в древних находках признаки человеческой культуры. К сожалению, во всех случаях «погребения» вполне могут быть объяснены естественными причинами (обвал, произошедший в пещере; повышенное содержание цветочной пыльцы в «могиле» из-за подстилки, используемой для ночлега и т. д.).
Теперь пришло время поговорить об орудиях неандертальцев.
Хотя неандертальцы использовали в том числе деревянные и костяные орудия, все же известность приобрели они благодаря своим каменным орудиям. И дерево, и камень подвергались старательной и весьма искусной обработке. Результатом такой обработки, например, были ручные рубила, которые начали делать еще в нижнем палеолите, т. е. за сотни тысяч лет до появления Homo sapiens. Напомним, что классические неандертальцы и гейдельбергцы жили в среднем палеолите (от 250 до 40 тыс. лет назад), так что орудия означенного типа появились задолго до них, еще во времена эректуса. Однако неандертальцы довели их до совершенства. Напомним, неандертальцы являются создателями так называемой мустьерской культуры, для которой характерна виртуозная и тщательная обработка каменных орудий. Разнообразие этих орудий поражает своим богатством. Это и рубила, и топоры, и наконечники для копий, и скребла, и шила, и пробойники, и сверла, и т. д. Все орудия неандертальцев произведены с большой тщательностью, для изготовления каждого требуется до нескольких тысяч операций (начиная производством отщепов от материнского каменного ядрища до корректировки заготовок и так называемой «штриховки» уже почти готовых изделий). Если так можно выразиться, неандертальцы довели культуру каменных орудий практически до своего апогея. Для дальнейшего развития должен был произойти качественный скачок, который, например, совершили кроманьонцы, изобретя лук и стрелы, метательные орудия типа боло, копья, предназначенные для метания, копьеметалку, полировку лезвий, и т. д… У неандертальцев этого качественного скачка не произошло, мустьерская культура на протяжении тысячелетий оставалась неизменной, точнее, она претерпевала очень медленную эволюцию в раз и навсегда заданном направлении. Увеличивалась тщательность изготовления каменных рубил, возникали новые модификации скребел, но ничего принципиально нового не появлялось.
Считается, что неандертальцы были искусными охотниками. Они охотились на довольно крупных травоядных, и даже на мамонтов. Тем не менее, большинство ученых склоняются к мнению, что метательных орудий у них не было: добычу убивали ударом коротких копий, о чем свидетельствуют следы развитых мышц на костях правой руки. Кроме того, короткие руки неандертальцев не способствовали развитию навыков в метании копий и иных предметов. Даже если бы они захотели сделать что-то подобное, вряд ли у них это получилось бы хорошо. Поэтому основная охота проходила, если так можно выразиться, в «ближнем бою». Подобный промысел был сопряжен с большими физическими нагрузками и немалым риском. Установлено, что неандертальцы устраивали ловушки на животных, проводили так называемую «загонную» охоту. Имеются находки, косвенно свидетельствующие о том, что неандертальцы заботились о пожилых членах сообщества, а также раненых на охоте соплеменниках. В частности, при анализе 36 скелетов неандертальцев, имеющих переломы, только у 11 результаты лечения перелома признаны неудовлетворительными. Из этого ряд ученых сделали вывод, что у первобытных людей имелись навыки лечить переломы. Впрочем, это же может говорить и о том, что «сложные» переломы практически всегда заканчивались смертью больного. Срастались только те, которые не были открытыми, не сопровождались смещениями и проч. Другими словами, «срастались» те, которые и не требовали внешнего вмешательства.
О заботе неандертальцев о своих раненых соплеменниках говорит находка в пещере Шанидар (Ирак), датируемая 50 тыс. лет назад. Она показала, что при обвале в гроте погибли двое калек, которые, очевидно, находились там одни, пока остальные члены группы охотились или занимались другими делами. У одного из них были свежие раны на ребрах, у другого – заживающий перелом черепа. Впрочем, данные интерпретации оспариваются другими учеными.
Нет также никаких бесспорных, подтвержденных данных на счет того, что неандертальцы хоронили умерших. То, что первоначально было интерпретировано как захоронение, вполне могло оказаться последствием обвала в пещере, а наличие повышенного содержания цветочной пыльцы в почве непосредственно возле скелета могло свидетельствовать об использовании травы (в которой, естественно, попадались цветы) в качестве подстилки для сна, а вовсе не в качестве «прощальных даров» умершему собрату. Очень много в литературе, посвященной неандертальцам, было уделено внимания и так называемому «культу черепов». Суть его состояла в том, что голова, отделенная от остального тела, захоранивалась отдельно в своего рода саркофагах, сложенных из камней. Логично было предположить, что подобные сооружения являлись объектом некого культа. Зловещий обычай вполне вписывался в картину, которая к середине XX века усилиями писателей и журналистов была нарисована о жизни неандертальцев – как о диких, злобных полуживотных, практикующих каннибализм. Общество с удовольствием воспринимало все эти басни, поскольку они возвышали обывателя в собственных глазах, наглядно показывая, насколько далеко он продвинулся по сравнению с неандертальцами по пути прогресса. Однако при более детальном изучении оказалось, что первоначальные описания раскопок страдали неточностями, что часто ученые, производящее их, выдавали желаемое за действительное. Практически все известные сооружения, представляющие собой объекты «культа черепов», могли образоваться (и, скорее всего, образовались) в результате естественных причин. Другими словами, никаких достоверных, бесспорных данных о том, что у неандертальцев имелись зачатки религиозных воззрений и искусств, на настоящий момент нет.
Впрочем, свидетельства каннибализма имеются в слишком большом количестве, чтобы ими можно было пренебречь. Скорее всего, неандертальцы, жившие в суровых условиях ледникового периода, довольно часто съедали своих умерших сородичей. Из более чем трех сотен найденных скелетов хорошо сохранилась только дюжина. Остальные имеют следы отделения мяса, в костях содержатся крошечные осколки кремневых орудий. Ничего необычного или особо ужасного в такой картине нет. Каннибализм – вполне рядовое явление и для Homo sapiens. В ряде первобытных племен он был широко распространен еще во второй половине XX века. А в экстремальных ситуациях (голод, война и т. п.) случаи каннибализма отмечались и среди наших вполне цивилизованных современников.
Все эти факты говорят лишь об одном – о крайнем практицизме неандертальцев, что, в принципе, не удивительно, если помнить о суровых условиях ледниковой эпохи, в которых им приходилось жить.
О причинах исчезновения неандертальцев спорят, но существует несколько наиболее распространенных версий.
1. Гибель из-за климатических изменений во время последнего (Вюрмского) оледенения: тяжелые условия жизни, падение рождаемости, высокая детская смертность;
2. Распространение болезней, в том числе занесённых из Африки современным видом людей и распространявшихся среди каннибалов;
3. Ассимиляция людьми современного типа. Как отмечалось выше, существует ряд доказательств, что у неандертальцев и кроманьонцев существовали «смешанные браки», потомство которых имело гибридные черты. В соответствии с современными представлениями о расселении человека из Африки, население всех континентов (кроме Африканского) происходит от небольшой группы людей, некогда перешедшей через Красное море на Аравийский полуостров. Их контакты с неандертальцами должны были происходить на берегах Персидского залива.
4. Вытеснение людьми современного типа. Кроманьонцы пришли в Европу около 40–50 тысяч лет назад, а через 28–30 тысяч лет неандертальцы полностью вымерли. Эти 20 тысяч лет сосуществования двух видов были периодом острой конкуренции за еду и другие ресурсы, победу в которой благодаря десятикратному численному превосходству и более интенсивному освоению земель одержали кроманьонцы.
Остается добавить, что во время Вюрмского оледенения (110-12 тыс. лет назад) Пиренейский полуостров был убежищем, где неандертальцы жили на протяжении еще нескольких тысячелетий после того, как вымерли в остальной Европе. Известны даже находки (впрочем, спорные) останков неандертальцев, относящиеся к историческому времени.
7
История человека современного типа – это ничтожно малый отрезок времени по сравнению со всей остальной историей эволюции гоминид. До этих пор мы говорили только о периоде в последние четыре миллиона лет, не касаясь периода от 7 до 4 миллионов лет назад. Тем не менее, это очень важный период, поскольку именно тогда произошло возникновение прямохождения и выход предков австралопитеков и Homo в саванну.
Однако на этом эволюционном отрезке мы сталкиваемся с гораздо большими трудностями. Если находки австралопитеков и предков Homo довольно редки, то находки окаменелых останков наших еще более отдаленных предков можно буквально сосчитать по пальцам. В силу недостаточности данных, восстановить эволюционные процессы, относящиеся к этому периоду, практически невозможно. Мы можем лишь гипотетически реконструировать некоторые черты многомиллионной эволюции и на их основании строить в большей или меньшей степени достоверные гипотезы.
Если говорить об ископаемых останках гоминид в целом, то можно сказать, что ученым в каком-то смысле повезло. Дело в том, что на юго-востоке Африки, как раз в том месте, где начинали свой эволюционный путь гоминиды, проходит большой тектонический разлом. Две плиты столкнулись, и одна из них «наползла» на другую, наклонившись и встав практически «торчком». Тектонический срез, таким образом, оказался на поверхности, представив на всеобщее обозрение весь «слоеный пирог», составленный из отложений разных периодов. Но это уникальное явление, которое в природе встречается не так часто. В остальных случаях интересующие нас окаменелости залегают на глубине от 700 метров до полутора километров (в зависимости от толщины слоев). К сожалению, скорость образования отложений такова, что пласт, относящийся, скажем, к третьему миллионолетию до нашей эры, будет залегать на такой глубине, что добраться до него будет возможно только с применением глубинного бурения. А для поиска редких артефактов, относящихся к эволюции гоминид (каковыми являются окаменелости), требуются значительные площади. Именно поэтому поиск окаменелостей производится в основном в местах трех типов: 1. Тектонические разломы естественного происхождения; 2. Горные разработки, где добываются полезные ископаемые (мрамор, щебень, песчаник, известняк, уголь и проч.); 3. Пещеры и прочие укрытия естественного происхождения, где скорость формирования отложений на порядки ниже, чем на открытой местности.
Нетрудно догадаться, что если глубина залегания пластов, относящихся к 3–4 миллионам лет назад, довольно значительна, то глубина залегания пластов, относящихся 7–8 миллионам лет назад будет примерно в 2 раза больше. Это составляет дополнительную трудность в поиске окаменелых останков наших более отдаленных предков. Впрочем, вполне возможно, в ближайшем будущем ученые порадуют нас новыми находками, поскольку в последнее три-четыре десятилетия интерес к подобного рода исследованиям очень сильно возрос.
8
Итак. Что же имеется на настоящий момент?
Для начала приведем расширенную до масштабов 35 млн. лет. таблицу эволюции высших приматов (рис. № 2).
Как видим, генеалогическое древо начинается родом древнейших ископаемых обезьян парапитеков (Parapithecus). Находки их отрывочны и недостаточны (фрагмент нижней челюсти с зубами). Жил парапитек предположительно около 35 млн. лет назад. Это были древесные обезьяны, питавшиеся фруктами. Строение зубов парапитека сочетает в себе как прогрессивные черты, так и общее сходство с зубами некоторых низших приматов, особенно долгопятов. Существует предположение, что парапитек – начальная стадия в эволюции человекообразных обезьян и человека. Впрочем, некоторые антропологи опровергают это, отрицая саму возможность отнесения парапитека к приматам.
Проплиопитеки (Propliopithecus) – следующая стадия. Жили они в нижнем олигоцене. Найдена только одна нижняя челюсть представителя этого рода. Её обнаружили в окрестностях города Эль-Файюм (Египет) еще в 1911 году вместе с нижней челюстью парапитека. Морфология зубов сближает этих обезьян с ныне живущими гиббонами, но они сильно уступают последним по размерам тела (в длину около 40 см). Обычно проплиопитеков считают предками плиопитеков и предположительно проконсула (камойяпитек).
Камойяпитек жил в Африке во второй половине олигоценового периода (прибл. 24,2-27,5 млн лет назад). Имеет значительное сходство с родом проконсул и был первоначально отнесен именно к нему, но потом на этот счет возникли споры. Камойяпитек известен исключительно по находкам зубов и челюстей. Сравнение зубов и челюстей камойяпитека, показало его схоство с, как уже говорилось, с проконсулом, а еще с моротопитеком. Однако, фрагментарность находок пока затрудняет решение вопроса о том, был ли камойяпитек одним из предков человека.
Далее следует моропитек. У моропитека (Morotopithecus bishopi), есть сильное сходство с афропитеком (Afropithecus), хотя тот жил несколько позднее. Моротопитек имел очень крупные, выступающие вперед челюсти с большими клыками, питался, как и его предки, плодами. Ростом был около 1,2 м и весил 40–50 кг. Радиометрическим методом моротопитек был датирован возрастом – 20,6 млн. лет. Если моротопитек и не является нашим предком, то похож он на общего предка человека, гиббоновых, орангутанов, горилл и шимпанзе, гораздо больше, чем проконсул (Proconsul). Дело в том, что у моротопитека, в отличие от четвероногих обезьян, часть поперечных отростков позвонков, к которым крепятся мышцы спины, развернута назад, что позволило ему, за счет тонуса мышц спины, хотя бы частично распрямить свою спину. Видимо, после моротопитека (как считает ряд ученых), предки человека уже никогда не были полностью четвероногими.
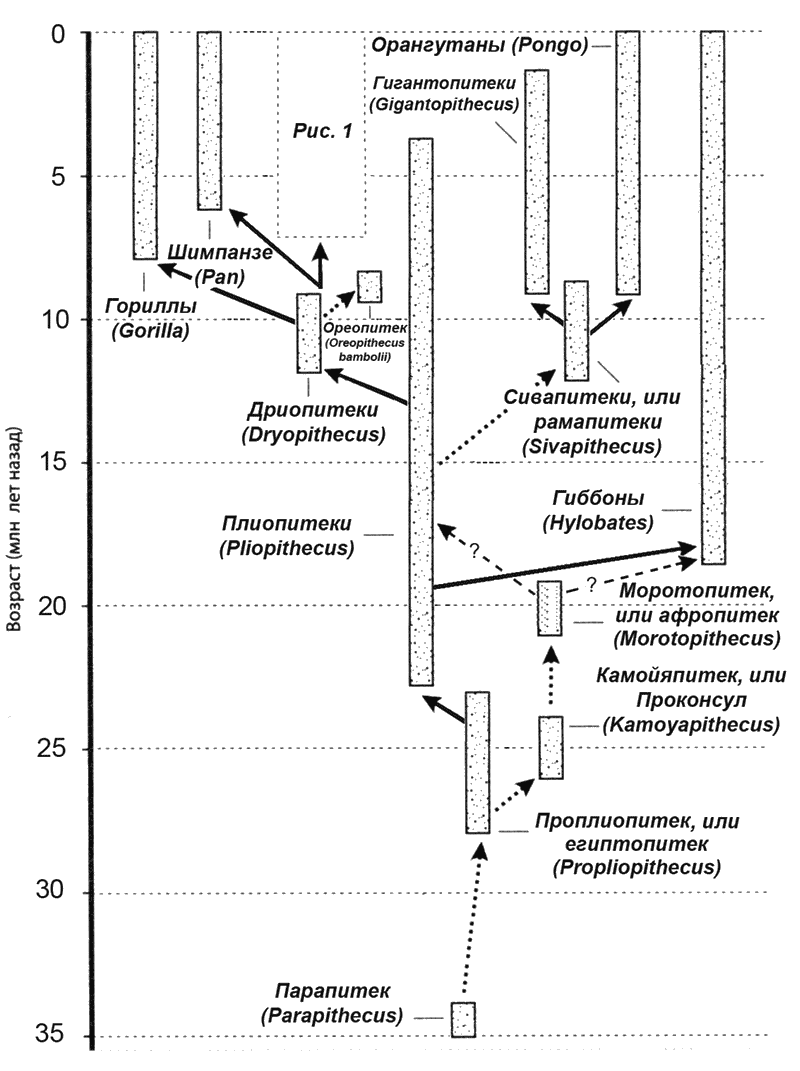
рисунок 2
Плиопитек – самый распространенный вид ископаемых обезьян, существовавший на протяжении почти 20 млн. лет (примерно с 23 до 3 млн. лет назад). Челюсть плиопитека впервые была найдена в 1837 г. во Франции, и это была вообще самая ранняя находка ископаемого антропоида. Как и на челюстях других антропоидов, подбородочный отдел у плиопитека отступает далеко назад. Клыки выдаются еще сильнее, чем у проплиопитека. В целом челюсть обладает столь явным сходством с челюстями гиббонов, что, несомненно, плиопитек (известный ныне по челюстям и зубам из Франции, Германии, Швейцарии и Монголии) – первый кандидат на роль их предка.
Плиопитеки населяли тропические леса, покрывавшие в те времена огромные области материков Европы, Азии и Африки. Изобилие пищи и сравнительно небольшое количество хищников на деревьях благоприятствовали их процветанию, да и благоденствию остальных видов обезьян.
В род Сивапитек (Sivapithecus) в настоящее время входит три вида, живших в период от 12,5 до 8,5 миллионов лет назад. С обнаружения первых ископаемых находок их отличало большое сходство с черепом орангутана, что подкрепляло теорию о том, что сивапитек был близким «родственником» орангутана. Женских особей этого рода долгое время ошибочно выделяли в род рамапитек. Наиболее крупный вид сивапитека Sivapithecus Parvada, вероятно, дал начало виду гигантопитеков.
Гигантопитеки (Gigantopithecus) обитали в юго-восточной Азии от 9 до полутора миллиона лет назад. По оценкам специалистов, гигантопитеки имели рост до 3 метров и весили от 300 до 550 кг, то есть были самыми крупными обезьянами всех времен. Считается, что питались гигантопитеки в основном бамбуком. Такой вывод делается из строения их зубов, которые демонстрируют приспособление к пережевыванию растительной пищи, а также того факта, что рядом с остатками гигантопитеков находили окаменелости панд, известных своей привычкой питаться исключительно побегами бамбука. Будучи похожи по строению тела на современных горилл, они, вероятно, передвигались на четырех конечностях.
На исходе плейстоцена гигантопитеки могли сосуществовать с людьми вида Homo erectus, которые начали тогда проникать в Южную и Восточную Азию из Африки через Ближний Восток.
Окончательно неизвестно, что именно было причиной вымирания гигантопитеков. Большинство исследователей считает, что среди главных причин были климатические изменения и конкуренция за источники питания со стороны других, более приспособленных видов – панд и людей, а также, возможно, изменения фауны, служащей источником их питания.
Помимо сивапитеков от плиопитеков произошли еще дриопитеки, вид, давший начало дошедшим до нас двум видов обезьян – гориллам и шимпанзе, а также линии, ведущей к австралопитекам и Homo.
Дриопитеки (Dryopithecus) известны по ряду остатков, найденных в Восточной Африке и Евразии. Жили они во времена миоцена, примерно 12-9 миллионов лет назад. Начав эволюционировать в южной части Восточно-Африканской рифтовой долины, предок дриопитека распространился по всему Африканскому континенту и Аравии, проник в Азию и Европу. Дриопитеки обитали на деревьях и питались, вероятно, ягодами и фруктами, так как найденные коренные зубы покрыты очень тонким слоем эмали. В длину он был 60 см, и, возможно, имел более длинные передние конечности, с помощью которых передвигался с ветки на ветку, как современные орангутаны и гиббоновые. Некоторые ученые полагают, что для дриопитеков был характерен стадный образ жизни. Некоторые авторы на основе особенностей скелета дриопитека высказывали предположения о его бипедализме (двуногости), что, однако, в дальнейшем не было подтверждено.
Более или менее определенно о прямохождении можно говорить относительно ориопитека (Oreopithecus bambolii), и именно этим примечателен в первую очередь данный вид. Жил ориопитек 9,5–8 млн. лет назад и происхождение свое, по всей видимости, ведет от дриопитека (хотя это многими оспаривается). Ореопитека называют также болотной обезьяной (Swamp Ape). Останки его были найдены в Италии, в Тоскане и на Сардинии, а также в Восточной Африке. Только в Италии найдены кости около 50 особей, что позволило хорошо изучить этот вид.
По размерам и подвижности ореопитеки были сходны с современными обезьянами. Весили они около 30–35 кг. Морда у них была сравнительно короткой, строение зубов – характерным для животных, питающихся листьями. Нижнюю часть лица они имели массивную, жевательный аппарат очень мощный, хотя зубы сравнительно мелкие. Клыки у них были не крупные, что многие авторы считают признаком отсутствия внутривидовой конкуренции и полового диморфизма.
Голова ориопитека имела округлую форму. Подобное строение черепа определило и особенности прикуса ориопитека, напоминающего прикус гоминид.
Но самое примечательное состоит в том, что этот примат практически перешел к прямохождению, хотя при ходьбе и придерживался руками за нависающие над головой ветви кустов и низких деревьев. Об этом вполне определенно свидетельствуют особенности строения рук, ног и позвоночника.