Часть I Параметры биогеографии
Чудище обло, озорно, огромно, стозевно и лаяй.
Глава 1 Введение в биогеографию
Хочешь обнять весь мир – купи глобус!
1.1. Предмет и задачи биогеографии
Старинная латинская мудрость гласит, что всякое знание начинается с названия. Название – биогеография – свидетельствует о дуалистичности этой науки, которая совмещает в себе биологические и географические аспекты. Она сосредоточена на такой важной стороне жизни, как необходимость пространства для её проявления. При этом внимание биогеографии к биологической и географической составляющим своей сущности в очень высокой степени сбалансировано. Начинается биогеография с хорошего знания живых объектов.
Мир живых существ на Земле очень разнообразен. Давно миновали времена, когда учёные-биологи пытались ограничить цифру разнообразия величиной 1 млн. видов. Ещё полвека назад эта цифра казалась недостижимо огромной. В течение последних 30 лет новые возможности науки и техники позволили поднять “планку” биоразнообразия до 1,5; потом 2, 5, 10, а потенциально – и до 80 млн. видов (Storck, 1988; Примак, 2002). Постепенно стало очевидным, что разнообразие видов является величиной, во-первых, непостоянной, а, во-вторых, бесконечной. Непостоянство заключается в следующем: новые виды животных, растений и протестов непрерывно возникают, а старые постепенно вымирают. Скорость обоих процессов, идущих то параллельно, то последовательно, меняется по ходу истории планеты на порядки величин. Бесконечность разнообразия форм жизни предопределена, с одной стороны, законами эволюции, а с другой – возможностями науки-систематики, призванной искать и расставлять по рангам признаки, различающие живые объекты. Опыт показывает, что число признаков, используемых в биосистематике, непрерывно растёт. Один исследователь, ради любопытства, насчитал у двух очень похожих жужелиц (рода Carabus) морфологические различия по 500 признакам и на этом остановился. Различия не кончились, но ему просто надоело их находить. А ведь, кроме морфологических признаков, систематики сегодня оперируют также признаками цитологическими, генетическими, иммунологическими и т. д. Их употребление необычайно расширило возможности био систематики, а, значит, и пределы биоразнообразия.
В такой ситуации остро встаёт вопрос, как познать бесконечный мир биоразнообразия, как упорядочить его восприятие человеческим мозгом? Этой цели служит всем хорошо знакомая универсальная познавательная операция, именуемая сравнением. Основополагающим принципом сравнения является корректность. Сравнивать корректно – значит сопоставлять предметы и явления между собой не как попало, а относительно единой стандартной логической системы критериев. К сожалению, корректность сравнения выдерживается не всегда.
Для получения хорошего результата при сравнении биологических объектов используют обычно три типа природных шкал.

Рис. 1. Эволюционное древо сравнения важнейших групп животных по родству (по: Николов, 1986).
Одной из них служит знаменитое филогенетическое древо жизни, придуманное и построенное в первом варианте великим естествоиспытателем Э. Геккелем. Оно представляет собой систему ветвей, соединённых друг с другом последовательно в определённом порядке. От основания древа к его вершине сложность строения живых существ растёт. Примерив к древу любое живое существо, можно определить, например, одноклеточное оно или многоклеточное, растение или животное. А также: злак или бобовое, насекомое или хелицеровое, жук или бабочка, бабочка-крапивница или бабочка-капустница и т. д. Чем более похожи между собой сравниваемые представители живого мира, тем ближе друг к другу располагаются они на филогенетическом древе (рис. 1). В последние десятилетия филогенетические схемы стали строиться с использованием не только фенетических, но и генетических не только качественных, но и количественных критериев. Примером служат так называемые кладограммы (от греч. кладос – ветвь), представляющие собой граф с дихотомически расходящимися ветвями. Каждая ветвь при этом удалена от исходной точки и других ветвей на расстояние, рассчитанное математически по сходству признаков сравниваемых таксонов (рис. 2). На базе такого сравнения друг относительно друга строится филогенетическая (иначе – генеалогическая) система организмов. Её изучают науки – ботаника, зоология, эволюционное учение.

Рис. 2. Кладограмма современных и некоторых ископаемых лошадей:
А – Equus asinus, Б – Е. burchelli, В – Е. caballus, Г – Е. grevvi, Д – Е. hemionus, Е. kiang, Е. onager, Е – Е. zebra, Ж – Е. quagga, М – Е. mauritanicus (плейстоцен, Европа), Т – Е. tabeti (плейстоцен, Алжир) (по: Эйзенманн, 1988).
Другая универсальная шкала сравнения – время, точнее тренды изменения во времени ключевых факторов среды обитания. Следуя трендам – циклическим и векторным, суточным и месячным, годичным и вековым – живые организмы перманентно мобилизуют свои адаптивные ресурсы из генетического резерва изменчивости, пока не исчерпают его полностью (рис. 3). Те, кто не поспевает за изменениями среды, вымирают, освобождая жизненное пространство более мобильным и генетически резистентным. Сравнение живых объектов относительно временных шкал позволяет различать организмы и их совокупности по ритмам онтогенеза, фенофазам, стадиям сукцессии, периодам геологической истории и т. п. Такое сравнение находится в компетенции палеонтологии, экологии, биономии.
Третий тип шкал сравнения задан структурой пространства земной поверхности. Условия жизни там меняются закономерно и градиентно вдоль сети параллелей и меридианов, от экватора к полюсам, от геометрических центров материков к океанским просторам, от подножия гор к их вершинам, от водораздела к местам аккумуляции водного стока и т. п. (рис. 4).
Именно сравнением биоразнообразия относительно пространственных шкал занимается биогеография, которая призвана решать три главные задачи:
1) анализ пространственных закономерностей планировки экологических условий на земной поверхности;
2) восприятие, осознание, анализ и прогноз картины распространения жизни на популяционно-видовом, синэкологическом и биоценотическом уровнях её организации;
3) использование выявленных закономерностей в качестве ключа к познанию истории Земли, эволюции живой природы и правил природопользования.
1.2. Истоки биогеографии
Биогеография ведёт свое начало от умения первобытного человека освоиться в пространстве с утилитарными целями. В мифах, легендах, религиозных преданиях (например, вавилонских сказаниях о Гильгамеше, библейских повествованиях о всемирном потопе, еврейских преданиях о скитаниях Моисея и других) сформулированы первые представления о биогеографической картине мира и месте человека в ней. Постепенно крепнущая уверенность в том, что без подобного знания людям на Земле не выжить, привела, в конце концов, к возникновению биогеографии.
В каждой уважающей себя науке на определённом этапе ее развития обязательно появляется собственный основоположник. Он необходим для того, чтобы, во-первых, заострить внимание на наличии основ; во-вторых, для возложения венка ответственности за всё уже содеянное этой наукой на конкретное лицо (желательно после его смерти, чтобы не отрёкся). Выбор основоположника в подавляющем большинстве случаев является делом тяжёлым! Тут всё имеет значение: разногласия по поводу того, что считать основами, личные качества претендентов, национальные претензии… Кроме того, нередко похожие мысли, идеи, обобщения, как назло, приходят в головы сразу нескольким лидерам того или иного научного направления. Казалось бы, выход прост – не скупиться на основоположников.
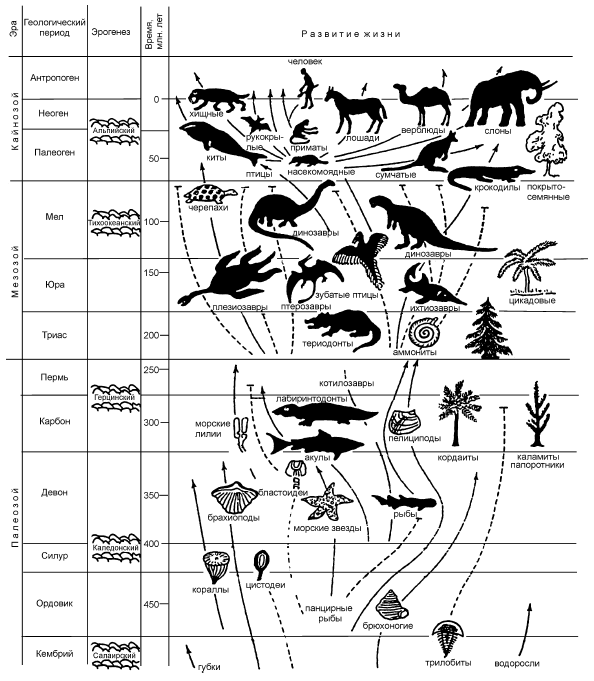
Рис. 3. Схема развития органической жизни относительно шкалы геологических периодов. Сплошные линии показывают, что группы организмов дожили до настоящего времени, пунктирные обозначают полностью вымершие группы.
Но тогда возникает реальная опасность каскадного эффекта на ярмарке научного тщеславия. На всякий случай, вошло в моду подбирать великих учёных попарно. Например: политологов – Маркса с Энгельсом, эволюционистов – Дарвина с Уоллесом, физиков – Фредерика Кюри с женой Марией Склодовской, почвоведов – Костычева с Докучаевым, генетиков – Уотсона с Криком и т. д. В этом плане биогеография составляет одно из счастливых исключений, так как в пару к Александру Гумбольдту поставить некого (рис. 5).


Рис. 4. Сравнение видов животных и растений вдоль шкалы высот в Кении (Африка) (по: Бигон и др., 1989) и на Алтае (Азия) (ориг.).
Некоторые историки науки (Наумов, 1969) предшественником А. Гумбольдта называют француза Ж. Бюффона, якобы основавшего “историческую” зоогеографию. Однако это не так! Бюффон был человеком безусловно, талантливым, но являлся больше гением интуиции, чем последовательным учёным. По мнению Ж. Кювье, его палеогеографические построения скорее угаданы, чем доказаны. Зато писателем Ж. Бюффон был замечательным, поэтому имел и до сих пор имеет много поклонников. Но он не подходит в качестве основоположника биогеографии в пару к Гумбольдту ни по времени, ни по сути.
1.2.1. Жизненный путь основоположника
А. Гумбольдт родился в Берлине 14 сентября 1769 г. и умер там же, прожив, дай Бог каждому – без малого 90 лет. Его путь к научному Олимпу был стремителен, а пребывание на вершине долгим. Ещё в нежном возрасте хрупкий мальчик увлёкся коллекционированием растений, насекомых и прочей, с точки зрения родителей, “дряни”, за что удостоился прозвища “маленький аптекарь”. Оно отнюдь не было лестным, ибо “аптекарями” тогда обзывали тех, кого сейчас преуспевающие господа дразнят “ботаниками”, “букашечниками”, “паганелями”. Самое интересное заключалось в том, что будущего великого естествоиспытателя угораздило родиться в семье прусских военных, да ещё и аристократов. Наследник был не просто Гумбольдтом, а носил имя: Александр Фридрих Вильгельм барон фон Гумбольдт. Он должен был служить, т. е. делать карьеру при Прусском дворе, а не пребывать в джунглях.
Однако судьбе было угодно распорядиться юным бароном иначе. С младых ногтей его увлекли как раз джунгли, но вначале только “джунгли учёности”. В 1787 г. он поступил в университет Франкфурта-на-Одере, а через два года – в Геттингенский университет. В Гамбургской Коммерческой Академии он изучает иностранные языки, а во Фрайбургской – горное дело. В 1792 г. он поступает на службу в Горное ведомство, но и там, помимо основной работы, продолжает заниматься самообразованием. Он исследует подземные газы и состав воздуха, проводит опыты над гальванизмом на собственной аристократической спине, интересуется химическими процессами растительной и животной жизни… А в 1794 г. его потянуло на философию, а затем обуяла неистовая страсть к овладению пространством. Он совершает многочисленные естественнонаучные экскурсии в Гарц, по Рейну, затем в Бельгию, Голландию, Францию, Англию… После смерти матери, продав родовые имения, что по тем временам было верхом безрассудства, молодой Гумбольдт отправляется в город Иену, где три месяца радуется общению с поэтами Шиллером и Гёте. В конце того же года он отправляется в Италию, а затем в Париж. В Париже знакомится с ботаником Э. Бонпланом, с которым и отправляется в 1799 г. в своё самое знаменитое путешествие. Они изъездили всю экваториальную полосу американского материка вдоль и поперёк. И это в те времена, когда в Южной Америке цивилизацией, по сути дела, и не пахло!
Бонплан, хотя и ездил с Гумбольдтом в одни и те же места, в основоположники биогеографии, увы, не попал! Видимо одной тяги к путешествиям для этого недостаточно. Нужны ещё энциклопедичность знаний, ну и, конечно, талант. А. Гумбольдту, который до этого уже был ботаником и зоологом, географом и геологом, физиологом и климатологом, этнографом и философом, стать биогеографом было легко. А уж таланта ему явно было не занимать.

Рис. 5. Основоположник биогеографии А. Гумбольдт.
После американского путешествия Гумбольдт в течение нескольких десятилетий прилежно обрабатывал собранные материалы и публиковал многочисленные учёные труды, неизменно приковывая к себе всеобщее внимание. В 1799–1804 гг. он издал самое объёмное своё произведение – “Путешествие в равноденственные области Нового Света” в тридцати томах. Пятой части этого суперфолианта хватило бы, чтобы считать Гумбольдта основоположником биогеографии. Благодаря этому научному подвигу Александр Гумбольдт безоговорочно и сразу занял главное место среди естествоиспытателей начала XIX века. Даже великий Гёте вынужден был признать, что “ему нет равного по образованности, живым знаниям и разносторонности, подобной которой он ещё не встречал”. Гёте уподобил Гумбольдта целой Академии! Такие титаны естествознания, как Кювье, Гей-Люссак, Латрейль, лишь обрабатывали привезённые Гумбольдтом материалы.
В 1810 г. ему был предложен пост министра народного просвещения Пруссии. Однако он решительно отказался. Впрочем, от отдельных ответственных поручений Двора Гумбольдт уклоняться не мог. Даже для такого знаменитого человека это было чревато всякими непредсказуемыми последствиями. Гумбольдту приходилось сопровождать прусского короля в его официальных поездках или служить послом в разных странах, выполняя деликатные представительские миссии. Таким образом, помимо прочих многочисленных профессий, А. Гумбольдт был ещё и прекрасным дипломатом. Без знания основ дипломатии, кстати, основоположником никакой науки не станешь, а уж такой многоликой, как биогеография, тем более.
Неистовый Гумбольдт, достигнув почтенного возраста – 60-летия – и имея большие чины, должен был, казалось угомониться, так как уже вдоволь напутешествовался. Однако не тут-то было. Зуд странствий по-прежнему манил и не давал покоя.
В 1829 г. Гумбольдт получил приглашение от царя Николая I снарядить научную экспедицию в азиатскую Россию. И престарелый, увенчанный всеми мыслимыми наградами и регалиями, немец не утерпел и отправился в очередную поездку. Царь, которого на Руси за репрессии против декабристов и создание профессиональной армии считают изощрённым злодеем и тупым солдафоном, не поскупился на науку. Экспедиция была великолепно оснащена. За семь месяцев были обследованы нижний и средний Урал, потом Алтай; затем через Семипалатинск, Омск, Оренбург отправились в Астрахань. И по Каспийскому морю через Донскую область участники экспедиции вернулись в Санкт-Петербург.
После этого путешествия А. Гумбольдт на 15 лет осел в Берлине, где занимался придворными обязанностями в светлое время суток, а по ночам писал свой “Космос” – очередной гениальный труд. Первый том вышел в 1845 г., четвёртый – в 1858, а пятый уже не вышел из-за смерти автора 6 мая 1859 г.
Великий немец оставил в наследство старому слуге небогатое имущество, а потомкам неоценимые теоретические изыскания, среди которых биогеографические исследования занимают одно из самых видных мест. Он описал территории и характер распространения многих видов растений и животных, обнаружил связь между их ареалами и климатом, сформировал экологическое направление биогеографии, сделал первую попытку выделения ботанико-географических областей. Гумбольдт установил, что распространение разных организмов (растительных, животных и человека) подчиняется общим единым правилам. Выявил глобальную закономерность увеличения видового богатства от полюсов к экватору и, наконец, открыл такую генеральную биогеографическую закономерность, как широтная зональность. Важнейшие принципы биогеографии изложены им в книгах: “Идеи о географии растений”, “Картины природы”, а также в уже названных интегральных трудах “Путешествие…” и “Космос”.
Основоположников наук зачастую изображают эдакими сухарями, людьми “не от мира сего”. К Александру Гумбольдту подобное ни в коей мере не относится. Ничто человеческое ему было не чуждо. Он был очень представительным мужчиной (см. рис. 5), имел репутацию салонного светского “льва”, славился умением очаровывать людей и с удовольствием этому предавался. Правда, жениться и обзавестись детьми – не успел. Однако любил…, любил беззаветно…, любил капризную даму, которая почти век изощрённо терзала его сердце, душу и ум. Её звали… Наука. Среди множества её прелестей, биогеографические явно пользовались у Гумбольдта особым вниманием. Таким образом, он стал на века основоположником науки, где “Биос” и “Гея”, “Хорос” и “Хронос” сплелись в причудливый, неразмотанный до сих пор клубок.
1.2.2. После Гумбольдта
С легкой руки маститого основоположника биогеография в дальнейшем получила “права гражданства” среди прочих почитаемых наук и даже стала модной. Благодаря этому, биогеографические идеи послужили стартовой площадкой в развитии многих других научных направлений. Например, учение о макроэволюции живой природы сложилось у англичан Чарльза Дарвина и Альфреда Уоллеса во второй половине XIX века только после того, как им удалось, путешествуя по свету, сравнить жизнь в пространстве.
Исходной позицией палеогеографии поначалу тоже служили закономерности распространения жизни. Ещё до Гумбольдта французу Жоржу Бюффону для объяснения размещения похожих форм животных на удалённых материках первому пришла в голову идея о некогда едином континенте, расколовшемся в дальнейшем на куски, которые постепенно оказались в разных сторонах. Причины данного явления в середине XVIII века, разумеется, были неизвестны. Однако Бюффону было легче вообразить себе плавающие материки, чем верблюдов, бизонов, тапиров и страусов, форсирующих океан. Полтора века спустя немец Альфред Вегенер, хоть и был весьма образованным для своего времени геофизиком и климатологом, формулируя свою теорию мобилизма земной поверхности, поначалу тоже отталкивался от биогеографии, пытаясь объяснить, каким образом близкие виды сухопутных животных и растений с общими эволюционными корнями оказались разделёнными тысячами километров воды.
Морская биология без биогеографической базы долго была весьма аморфной областью знаний. Лишь в середине XIX века англичанин Эдвардс Форбс и одновременно американец Джеймс Дана разработали биогеографическую систему вертикальных и горизонтальных зон океана. Появившийся пространственный порядок позволил Дж. Дана ввести в морскую биологию количественный анализ. Это дало гидробиологии изрядное преимущество перед биологией наземной, ощущаемое до сих пор. На этой базе столетие спустя русский учёный Лев Александрович Зенкевич сформулировал учение о продуктивности и биологической структуре океана, в основе которого тоже лежат биогеографические закономерности.
Представления о биогеографической широтной зональности, идущие от А. Гумбольдта, привели россиянина Василия Васильевича Докучаева в начале прошлого столетия к созданию генетического почвоведения. А русских ботаников Гавриила Ивановича Танфильева (автора первой сводки по географии растительности России), Андрея Николаевича Краснова (первым обозначившего флоры России), Василия Васильевича Алёхина (автора первого геоботанического обзора) – к идеям геоботаники.
Один из последних учёных-энциклопедистов XX века Лев Семенович Берг, отталкиваясь от биогеографии, к середине столетия сформировал учение о ландшафтах.
Без биогеографической основы трудно представить себе возникновение и развитие микроэволюционных теорий видообразования Эрнста Майра, Джорджа Симпсона и других в XX веке. Кладистический подход к систематике биологических форм, разработанный немцем Вилли Хеннигом ещё в 30-е годы XX века, так и считали бы схоластическим, если бы Леон Круаза, Д. Нельсон, Норман Платник не подвели под него биогеографическую базу. С этого времени кладистика стала теорией.
Успешно оплодотворяя другие науки, биогеография при этом оставалась самодостаточной и бурно прогрессировала. Среди многочисленных последователей А. Гумбольдта непосредственно на поприще биогеографии следует выделить А Гризебаха, издавшего в 1827 г. мощную сводку “Растительность Земного шара”, где в сравнительном ключе учтено и проанализировано распределение растительности в соответствии с климатом и высотой места. В России Николай Алексеевич Северцов (1827–1889) превратил зоогеографию из описательной науки в аналитическую, используя сравнительно-географический метод для выделения фаун в их связи с климатом и почвой. Одним из первых стал использовать биогеографическое районирование в орнитологии Михаил Александрович Мензбир (1885–1935). Историю биогеографии украшают также другие славные имена. Ссылки на них будут часто встречаться на страницах этой книги.
Некоторое представление об уровне развития биогеографии к концу XX столетия даёт библиографический анализ, проделанный известным геоботаником и биогеографом Анатолием Георгиевичем Вороновым (1980). По его данным общее число изданных в мире работ по биогеографии составляло в 1975–1978 гг. – 28430. За истекшее после этого время цифра выросла приблизительно в полтора раза.
Таким образом, истоки биогеографии после А. Гумбольдта не иссякли, а превратились в обильный и разветвлённый информационный поток. Он свидетельствует об усилении интереса людей к правилам дифференциации земной поверхности и населяющей её жизни. Происходит это не вопреки, а наряду с ростом глобализации представлений о Земле. При этом стоит иметь в виду, что глобализация имеет пределы, заданные размерами планеты, а дифференциация практически неисчерпаема особенно с учётом динамики среды. Поэтому, до тех пор, пока есть жизнь на Земле, которая продолжает развиваться, организмы будут “кроить” пространство, деля его каждый в свою пользу…
1.3. Место биогеографии среди других наук, её структура
Биогеография не стоит среди других наук особняком, а существует на стыке родительских областей знания: биологии и географии. Из-за этого она имеет сложную структуру, которую можно представить в виде знаменитого кубика Рубика, чьи грани соответствуют определённым разделам биогеографии (рис. 6).
С позиций естественной классификации организмов в рамках биогеографии существуют: география растений (фитогеография), география животных (зоогеография), география человека (антропогеография) и география протистов (т. е. одноклеточных организмов – протистогеография). Каждый из этих разделов имеет свои оригинальные подходы к изучению распространения организмов, исходя из специфики их устройства. Например, распространение автотрофов-растений зависит, прежде всего, от распределения ряда абиотических факторов (света, тепла, влаги), а гетеротрофов-животных в большей мере – от размещения трофических партнёров. Благодаря предельно простой и потому универсальной для всех условий организации, объекты протистогеографии имеют почти неограниченные возможности для распространения. Люди тоже распространены повсеместно, благодаря не то обилию разума, не то его недостатку. Тем не менее, они представлены несколькими хорошо выраженными морфологическими расами, тяготеющими каждая к определённой части земного пространства.

Рис. 6. Инфрастуктура биогеографии (ориг.).
С позиций физического состояния среды обитания различают биогеографию суши (сухопутную биогеографию), биогеографию океана (морскую биогеографию), биогеографию внутриконтинентальных водоёмов (т. е. клочков воды, окружённых со всех сторон сухопутьем) и, наконец, биогеографию островов (т. е. “клочков” суши, окружённых со всех сторон морем). Для суши характерны очень протяжённые и контрастные хорологические градиенты экологических факторов. В морской среде контрастность пространственных градиентов в значительной степени нивелируется физическими особенностями жидкого субстрата. На суше изменения экологических факторов в пространстве задаются в большой мере перемещением воздушных масс, а в море – водных течений. Островная биогеография вынуждена в равной мере апеллировать к факторам как сухопутным, так и морским. Кроме того, решающее значение для распространения организмов приобретают абсолютные размеры острова, его удалённость от других островов и ближайшего материка.
С позиций уровня взаимодействия живых существ между собой и с абиотической средой в структуре биогеографии выделяют географию биологических таксонов (ареалогию), географию флор, фаун, биот (флористику, фаунистику), географию экосистем и, наконец, географию ноосистем (от гр. “nous” – разум), т. е. систем жизнеобеспечения, созданных человеческим интеллектом заново или с использованием диких природных элементов. Ареалогия исследует пределы, форму, протяжённость территории распространения отдельных таксонов биоты. Задачей ареалогии является выделение, описание, объяснение и типологизация (выделение типов) ареалов, но без объяснения функциональных связей обладателей этих ареалов. Флористика и фаунистика занимаются выявлением и классификацией флор и фаун, т. е. совокупностей таксонов, имеющих ареалы со сходными параметрами. Характер уживаемости видов в рамках одной флоры или фауны остаётся за скобками биогеографического анализа. Наконец, географию экосистем, в отличие от экологии, интересуют не сами по себе функциональные взаимоотношения компонентов экосистемы, а закономерности размещения отдельных функциональных параметров или целых сообществ, биоценозов и биогеоценозов в пространстве.
География ноосистем специализируется на изучении обязательных зависимостей творений человеческого разума от существующей инфраструктуры земного пространства.
1.4. Основные теоретические позиции современной биогеографии
В современной биогеографии сосуществуют, по крайней мере, четыре теоретических направления.
Первое – это дисперсионная биогеография. Она базируется на дарвиновской концепции монофилетического происхождения таксонов филогенетической системы (каждый – из одного корня). Любой таксон имеет своё характерное время и место происхождения. То и другое устанавливается путем разделения и расстановки по древности рецентных, т. е. ныне живущих, и ископаемых представителей того или иного таксона. На этой основе реконструируются траектории его расселения по планете.
Особенности географического распространения таксона с позиций обсуждаемой теории предопределены адаптациями к перемещению, характерными для его представителей. Поскольку транспортные возможности у организмов из разных таксонов сильно различаются, то равномерного распределения жизни по земной поверхности не получается. В разных её частях складываются неодинаковые совокупности живых форм, которые можно лишь формально называть фаунами, флорами и т. д., ограничивая их содержание инвентарным списком видов, родов и т. д. Целью дисперсионной биогеографии служит описание, объяснение и классификация географических ареалов как можно большего числа таксонов, в перспективе желательно всех (рис. 7).


Рис. 7. Сравнение ареалов – основа дисперсионной биогеографии. По: Городков, 1986.
Императивом дисперсионной биогеографии является убеждение, что биогеографическую картину организмы формируют случайным образом, оперируя лишь собственными возможностями расселения, при пассивной роли среды.
Другим очень популярным теоретическим направлением биогеографии является флористико-фаунистическое. В противовес предыдущему, оно акцентирует внимание на формировании ограниченного набора очагов жизни, чётко ориентированных в географическом пространстве. В каждом из них исторически давно и непрерывно генерируются и интегрируются в совместимые и специфические совокупности таксоны животных и растений с ареалами более или менее одинаковой протяжённости, формы и инфраструктуры. Именно их следует именовать флорами и фаунами. Из этих очагов происходит дальнейшее расселение организмов. Выявление флор и фаун “де-факто” происходит не только путём статистического сравнения списков видов, но и выделения руководящих таксонов, типичных для разных очагов. Не менее важным критерием служит предварительное ограничение территорий и их разделение по специфике условий жизни. Эта специфика предопределена наличием физических преград (барьеров), препятствующих как вселению, так и выселению биоты с определённых территорий. Барьеры, мешая свободному распространению организмов даже с хорошими расселительными возможностями, создают режим изоляции внутри ограниченного пространства и способствуют там естественному отбору специфических форм жизни. Благодаря барьерам, реальные ареалы многих таксонов значительно меньше потенциальных. Из-за неоднородности экологического фона в пространстве у многих таксонов случаются разрывы ареалов. У истоков этих гипотез, объясняющих данные постулаты, стоял А. Уоллес (гипотеза оттеснённых реликтов и другие). Таким образом, суммарная биогеографическая картина мира по флористико-фаунистической теории складывается под влиянием не случайного, как в дисперсионной биогеографии, а детерминированного распределения и комбинирования таксонов, адекватно инфраструктуре пространства, заданной априори. Стандартная матрица пространства “диктует” одним и тем же таксонам совершенно разный порядок распределения в разных местах географической арены или одинаковое распределение разных таксонов в определённом месте часто вопреки их биологическому потенциалу к расселению. В противовес дисперсионной биогеографии – флористико-фаунистическую биогеографию можно ещё назвать интеграционной. А её императивом становится признание того, что биогеографическая картина мира является следствием не только биологического потенциала организмов к расселению, но и детерминированности географической арены. Её структура диктует порядок интеграции жизни в устойчивые совокупности таксонов флоры, фауны и т. и., характерные по составу (рис. 8).

Рис. 8А. Схема расположения центров и путей распространения флор (по: Алёхин, Кудряшев, Говорухин, 1961) в Евразии:
Европейско-Сибирские лесные флоры: 1 – ледниковые убежища жизни и центры распространения флоры широколиственных и смешанных лесов и таёжной флоры; 2 – послеледниковые пути расселения флоры широколиственных лесов; 3 – пути расселения флоры хвойно-широколиственных лесов; 4 – скандинавские послеледниковые пути расселения флор еловых и сосновых лесов; 5 – пути послеледникового расселения таёжной флоры из Алтае-Саянского и Южно-Уральского убежищ жизни; 6 – расселение лесной флоры Берингии; 7 – пути расселения флоры светлохвойных (б.ч. лиственничных) и смешанных лесов Восточной Сибири из Северного Китая в третичном и четвертичном периодах; 8 – древний третичный центр происхождения флоры сибирских и американских хвойных лесов; Восточно-Азиатские и Средиземноморские лесные флоры: 9 – области реликтовых флор (третичных) листопадных и смешанных лесов Азии; 10 – плейстоценовые пути расселения флоры азиатских широколиственных лесов (Алтае-Саянский путь); 11 – северные (сибирские и урало-украинские) пути расселения третичных флор широколиственных лесов; 12 – флоры третичных субтропических вечнозелёных лесов Китайско-Японской подобласти и их реликты на Кавказе; 13 – флора Западной Средиземноморской подобласти; 14 – пути расселения флоры вечнозелёных и листопадных лесов из Юга Восточной Азии в Средиземноморскую подобласть в третичном периоде (Гималайский путь).

Рис. 8Б. Схема расположения центров и путей распространения фаун (по: Мекаев, 1987) в Евразии:
1 – ледниковые рефугиумы флоры широколиственных, смешанных и таёжных лесов; 2 – пути расселения флоры широколиственных лесов в постледниковье; 3 – пути расселения флоры хвойно-широколиственных лесов; 4 – пути расселения флоры хвойных лесов; 5 – области реликтовых флор третичного времени; 6 – фаунистические центры; 7 – пути расселения фаун.
Третье направление в биогеографии – теория викарианса (мобилизма). Она имеет в основе постулат, который гласит, что схемы филогенетического (эволюционного) порядка замещения таксонов и исторической последовательности трансформации структуры географической арены строго согласованы. Наиболее ярким примером мобилистической биогеографии является согласование эволюционных схем развития жизни с рассредоточением фрагментов мезозойского суперконтинента Пангея в ходе геологической истории планеты, рисуемое геофизиками (Еськов, 1984; Кафанов, Кудряшов, 2000). В соответствии с канонами обсуждаемой теории, биологические таксоны сначала расставляются с помощью кладистического анализа по степени отклонения от исходной формы. Затем по аналогии с кладограммами систематиков строятся биогеографические кладограммы, показывающие не только современную картину распространения какого-либо таксона, но и историческую последовательность её формирования (рис. 9).
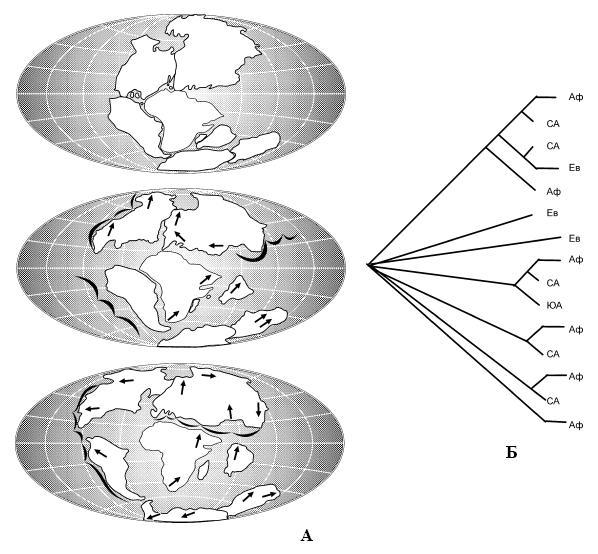
Рис. 9.
А – схема изменений местоположения континентов (по: А. Вегенеру из: Клечек, Якеш, 1985); Б – географические кладограммы лошадей по современным и ископаемым видам (по: Биосфера, 1988):
Аф – Африка, Ев – Евразия, СА – Северная Америка, ЮА – Южная Америка.
В качестве краеугольных камней викариантной (мобилистической или кладистической) теории рассматриваются следующие постулаты:
1) если таксон имеет ареал, разорванный какой-нибудь преградой, то его предки расселялись тогда, когда этой преграды ещё не было;
2) степень родства двух ныне изолированных контингентов таксона соответствует времени их изоляции, т. е. наиболее близкородственные из них представлены на наиболее поздно разошедшихся фрагментах некогда единого ареала;
3) более примитивные представители таксона всегда располагаются географически ближе к первичному центру происхождения, чем более продвинутые (Красилов, 1977).
Согласно императиву мобилистической (викариантной) биогеографии, современная биогеографическая картина составляет только преходящий эпизод в перманентной череде сочетаний биологических видов, замещающих друг друга в ходе эволюции, адекватно порядку изменения структуры географической арены. Благодаря неодинаковой скорости изменений в разных её частях единовременно, но разобщённо в пространстве сосуществуют серии заместивших друг друга таксонов, что обеспечивает гетерогенность флор и фаун. В какой-то мере это явление напоминает смену актёров на одну и ту же роль в киносериале по ходу изменения конъюктуры актерского рынка в разные периоды съёмок. Зритель же вынужден мириться со сменой лиц персонажей от серии к серии, утешаясь неизменностью сюжетной линии (рис. 9).
Четвёртое направление биогеографии опирается на теорию динамического равновесия, сконструированную американскими учёными Мак-Артуром и Уилсоном (1967) для нужд биогеографии островов. Суть теории заключается в том, что количество видов, населяющих остров, определяется равновесием между иммиграцией и вымиранием. Сколько видов вымирает – столько замещается за счёт вселения. В конечном итоге общее число видов на острове определённой площади и удаления от очага иммиграции остаётся приблизительно одинаковым (рис. 10). Если острова разной площади расположены на разном расстоянии от источника колонизации, то увеличение расстояния уменьшает скорость иммиграции, а увеличение площади острова – уменьшает скорость вымирания. Зависящее от площади острова равновесное число видов изменяется при этом по степенному закону. Поскольку островной эффект, в принципе, имеет место не только в океане, но и на суше (озёра как острова воды среди сухопутья, пятна степей среди тайги), то применение теории равновесия выходит за рамки морской биогеографии, её породившей.

Рис. 10. Теория равновесия Мак-Артура и Уилсона (по: McArthur, Wilson, 1967 из: Бигон и др., 1989):
А – зависимость скорости иммиграции от числа обосновавшихся видов для крупных и мелких островов, лежащих вблизи и вдали от континента; Б – зависимость скорости вымирания от числа обосновавшихся на острове видов для крупных и мелких островов; В – баланс между вымиранием для крупных и мелких и для близких и удалённых от континента островов. В каждом случае S – обозначает равновесное видовое богатство.
Императивом экологической биогеографии становится уверенность, что замещение таксонов, как эволюционное изменение форм путём естественного отбора, так и комбинаторное заимствование подходящих форм в пространстве из имеющегося по соседству ассортимента, происходит в любом месте географической арены обязательно под контролем конкретных экосистем, где все элементы скованы, как каторжники, единой цепью круговорота веществ и энергии. Даже при появлении новой, замечательной по своим потенциям формы жизни, её внедрение в какое-либо местообитание возможно лишь с “разрешения” экосистемы при наличии подходящей свободной экологической ниши.
1.5. Методы биогеографии
Несмотря на отмеченную специфику, все отрасли биогеографии роднит главное: все они изучают живые системы любого уровня организации с единых хорологических позиций, с помощью сравнительно-географического метода на базе количественных и качественных учётов встречаемости организмов в прошлом или настоящем.
Являясь наукой пограничной, биогеография опирается на главный метод и широко пользуется результатами исследований соседних отраслей знания. Уже было сказано о значении кладистического анализа, заимствованного из биосистематики. Находят применение кариологические методы. Широко используется изучение макроскопических остатков и микрофоссилий (игл, раковин фораминифер, спор, пыльцы растений). Переносящаяся в атмосфере пыльца может оседать на поверхности субстратов, благоприятных для её фоссилизации, при быстром погребении в условиях, способствующих её сохранению (на торфяниках, днищах озёр, в некоторых почвах). Определение и подсчёт зёрен пыльцы в образце, взятом через много лет, даёт возможность воссоздать средний состав и направление пыльцевого “дождя” в ту или иную эпоху, представить себе палеопейзаж и сравнить его с современным.
Широко заимствуются методы палеогеографии. С их помощью можно более или менее точно воссоздать характер поверхности Земного шара в далёком прошлом и представить себе пути миграции организмов. На этот счёт существует, например, палеогеографическая теория межконтинентальных мостов, которые временно связывали между собой крупные острова или целые континенты. Примером современных континентальных мостов, широко используемых биотой для расселения и расширения первоначальных ареалов, служат: Панамский перешеек между Северной и Южной Америками, Суэцкий перешеек между Африкой и Азией. Сравнительно недавно существовали и определяли миграции биоты Берингийский мост между Чукоткой и Аляской, перешеек Па-де-Кале между континентальной Европой и Британскими островами; мосты, соединявшие ещё в плиоцене юго-восток континента Азия и острова Индонезии. Очень продуктивна в биогеографии палеогеографическая теория дрейфа континентов (А. Вегенер) или, в свете новых данных, теория тектоники литосферных плит. Её суть, вкратце, сводится к тому, что несколько плотных и вечных кристаллических платформ “плавают” в поверхностном слое земной мантии (астеносфере) под влиянием конвективных течений магмы. Сталкиваясь и расходясь неоднократно в ходе истории планеты, эти плиты создают на земной поверхности в каждый геологический период своеобразную мозаику суши и моря, континентов и островов, горных и равнинных территорий, предопределяя на миллионы лет биогеографические закономерности.
Пользуется биогеография также методами палеоклиматологии в сочетании с методами геоморфологии. Речь идёт о реконструкции древних климатов, которые запечатлелись в особых формах рельефа, характере геологических отложений и почв. Например, такие формы рельефа, как морены, цирки, бараньи лбы указывают на оледенение и показывают границы соответствующих климатов, а, стало быть, и биогеографические границы. Качественными индикаторами климатов прошлого могут служить горные породы. Только в аридном климате образуются доломиты, ангидриты, гипсы, калийная и каменная соли, осаждающиеся из растворов в условиях сильного испарения. Индикатором жаркого сухого климата служат карбонатные красно цветы. Об аридном климате свидетельствуют лёссы и лёссовидные породы. Наоборот, только в гумидных условиях могли накапливаться каменные угли, бескарбонатные красноцветы, белые глины (каолины), алюминиевые руды (бокситы). Тёплому климату свойственны эвапориты, морские известняки. Свидетелями холодного климата служат так называемые тиллиты – не слоистые и несортированные конгломераты валунов с глинами или мергелями, рассматриваемые как остатки древних ледниковых морен.
Пригодными в биогеографии являются и геохимические методы. Например, по соотношению изотопов кислорода О16 и О18 в карбонатах можно судить о пале-температурах. Обогащение изотопом О18 растёт с понижением температуры, а О16 – с повышением.
Вопросы для самопроверки:
1. Чем отличаются науки биологического и географического циклов?
2. Назовите основные структурные подразделения биогеографии. Каковы принципы их выделения?
3. Какие теоретические направления биогеографии Вы знаете? В чём их отличия друг от друга?
4. Кто из учёных, когда и за что заслужил право именоваться “основоположником” биогеографии?
Глава 2 Проблемы биогеографии
Если все думают одинаково, значит никто особенно и не думает.
С тактической точки зрения посвящать вторую главу книги обсуждению проблем, стоящих перед пропагандируемой наукой, до того, как изложены её основные положения, рискованно. Однако с позиций стратегии познания суть любой науки заключается именно в решении проблем. Если проблем нет, то наука превращается в рутинное ремесло, ну а статус ремесленника вряд ли какому-либо ученому польстит. Поэтому, коль скоро, наличие проблем украшает науку, как боевые раны – мужчину, то лучше их не скрывать, а употребить для увеличения обаяния биогеографии.
2.1. Проблема научных границ от Уоллеса до наших дней
В любой науке, базирующейся на границе нескольких областей знаний, всегда есть опасность “сползти” в сторону одного из родительских направлений и потерять самобытность предмета и задач исследования. Такие проблемы есть и в биогеографии.
Первый звонок прозвучал ещё в середине XIX столетия в период формирования эволюционной теории. Искушение биологией было так велико, что два титана научной мысли Чарльз Дарвин и Альфред Уоллес, будучи поначалу добропорядочными биогеографами, так увлеклись изменчивостью, наследственностью и естественным отбором живых форм, что едва не превратили биогеографию в “золушку” биологии (рис. 11).

Рис. 11. Чарльз Дарвин и Альфред Уоллес.
Биографы А. Уоллеса рассматривают его жизненный путь обязательно в контексте судьбы Ч. Дарвина, называя их, как правило, соавторами эволюционной теории. Делается это, надо полагать, с целью подчеркнуть благородство Дарвина, выпускника привилегированного Кембриджа, обладателя высоких научных титулов, автора толстенных манускриптов об эволюции видов, но… согласившегося считать соавтором человека без общепринятого образования и определённой профессии, изложившего свои соображения по естественному отбору в небольшой статье, написанной не в солидном кабинете с библиотекой, а на диком острове под пальмой.
Между тем А. Уоллес по своей эрудиции в вопросах эволюции, а также по благородству души ничуть не уступал Дарвину. В разгар выяснения истины с приоритетом открытия естественного отбора Уоллес взял да и выпустил свою книгу “Дарвинизм”, определив, таким образом, авторство и дав тем самым название учению об эволюции. Впрочем, жертвуя первенством в области эволюционной теории в пользу соперника, Уоллес следом за первой издал вторую свою книгу о географическом распространении видов животных. Она получилась увлекательной и умной, уникальной по богатству материалов. Затем вышла ещё одна, посвящённая животному миру островов. Использованные в них обширнейшие и интереснейшие факты, закономерности, теоретические позиции, термины и понятия используются и сегодня. Увы, ответной любезности Дарвина в виде книги “Уоллизм” не последовало. Уоллес отличался от Дарвина не только ориентацией в науке, но и витиеватыми перипетиями жизненной судьбы.
Дарвин был человеком благополучным во всех отношениях. Ему досталась первоклассная наследственность от своего знаменитого деда – Эразма. Род Дарвинов был не из самых бедных. Чарльз получил блестящее образование, обучаясь в Эдинбургском и Кембриджском университетах. Дарвину потрясающе везло. Ничем не примечательным юнцом он каким-то чудом попал на корабль “Бигль”, который отправлялся в кругосветное путешествие. Если бы капитан Фиц-Рой заупрямился, то не было бы знаменитого Дарвина. А надо отметить, что он сразу невзлюбил нового натуралиста. В те времена мало было собрать коллекции, их ещё надо было вывезти на утлых кораблях. Дарвину и тут сильно повезло. Он всё привёз в Лондон в целости и сохранности. На этих собранных материалах и была построена теория естественного отбора. Свои книги Дарвин писал в идеальных условиях уютного кабинета с обширной библиотекой в собственном доме недалеко от Лондона, окружённый заботами любящей жены, сына Френсиса в качестве секретаря, многочисленной угодливой челяди.
Уоллес, не в пример Дарвину, попал в науку путями Господу неисповедимыми. В юности он собирался стать… часовщиком. Кто знает, может быть, это была неосознанная тяга к познанию времени как категории, относительно которой он потом сравнивал организмы? Научившись разбирать часы, он ушёл… в землемеры. Быть может, это тоже было подспудное стремление к пространственной оценке вещей и явлений. Работая в полях, он невольно наблюдал и собирал растения под ногами, таким образом, приобщаясь к биологии. Через некоторое время он сменил профессию землемера на должность учителя. Не преуспев и в этом, стал строить железнодорожные будки для обходчиков. Затем торговал насекомыми. Эта стезя завела его в джунгли Южной Америки. Вот там-то он по-настоящему страстно увлёкся устройством природы. Как многие гении, Уоллес ради науки мог пожертвовать всем. Когда в джунглях Амазонии он оказался перед сложным выбором: везти опасно больного брата в город или продолжить работу, предоставив умирающего провидению, то Уоллес, не задумываясь, выбрал второе. Брат умер, собранные Альфредом коллекции погибли на обратном пути, но в результате Уоллес всё-таки стал натуралистом. Несколько лет спустя, уже в Малайзии, он заболел жёлтой лихорадкой, но выжил. Более того, именно во время болезни Альфреду, когда он находился между жизнью и смертью, а тяжёлое состояние его усугубляли ещё и муки совести по поводу судьбы брата, умершего от той же болезни, пришли в голову самые замечательные эволюционные и биогеографические мысли и идеи,
Когда Уоллес вернулся домой в Англию, то выгодно продал свои кллекции, содержащие более 15 тысяч бабочек, 83 тысячи жуков, 8 тысяч птиц. На вырученные деньги он далее жил безбедно, писал свои статьи и книги. Последние 20 лет его жизни обычными для учёного не назовёшь. Он увлёкся спиритизмом. Нет, не от скуки и не “с глузда съехавши”, как нередко намекают биографы. Уоллеса, кроме изменения организмов во времени (эволюции), их изменения в пространстве (биогеографии), заинтересовали поиски механизма формирования умственных и нравственных способностей человека, которые он считал независимыми от естественного отбора. Спиритизм понадобился, чтобы вызвать к себе на суд духов Наполеона, Спинозы и, главное, умершего к тому времени Дарвина. Если отвлечься от несолидного спиритизма, то Уоллес был, по сути дела, первым человеком, который пытался создать синтетическое представление о жизни как интегральной функции времени, пространства и интеллекта. Однако дальше спиритизма это дело у Уоллеса не пошло. Стройное учение о ноосфере (сфере человеческого разума) как особой оболочке Земли создал через несколько десятилетий В.И. Вернадский. А Уоллесу досталась в истории науки роль дублёра Дарвина. На самом деле это совсем не так! Дарвин, поначалу вкусивший от биогеографии, далее навсегда увлёкся процессом изменения живых форм во времени. Уоллес же, переболев эволюционной лихорадкой, пошёл по пути познания и продуктивно использовал эволюционные биологические идеи для объяснения закономерностей изменения жизни в пространстве. Таким образом, он, в сущности, оказался “спасителем биогеографии” от “биологического ига”, пожертвовав ради этого славой основателя теории эволюции.
Попытки “утянуть” биогеографию под крышу какого-нибудь из родительских направлений продолжались и после Уоллеса. С превращением А. Вегенером идеи мобилизма (т. е. подвижности земной поверхности) из “бюффонады” в стройную обоснованную теорию, биогеографию хотели вернуть в родовое гнездо классической географии. Сторонникам такой процедуры казалось, что детерминированная структура земной поверхности априори диктует биоте, как распределяться в пространстве. Однако, оправившись от первого шока, биогеографы возвратились к убеждению, что даже при большом удалении материков друг от друга, при полной физической изоляции, характер распространения биологических таксонов определяется, кроме географии, ещё и генетическим родством эволюционных предков.
С того момента, как Эрнст Геккель провозгласил науку “о домохозяйстве организмов”, в научном мире зрело желание подложить “яйцо” биогеографии под гостеприимную наседку – экологию. Апофеозом этих экспериментов стала попытка американского учёного Роберта Мак-Артура, опять-таки на пару с Эдвардом Уилсоном, заново сформулировать биогеографию языком принципов популяционной экологии и генетики. В результате они оказались, по их словам, “не в состоянии увидеть какие-либо реальные различия между биогеографией и экологией”. Во многих учебниках и руководствах по биогеографии 1960-80-х гг. и даже последних лет чисто экологические разделы по объёму текста и информации превысили биогеографические (Леме, 1976; Воронов, 1999; Петров, 2000). Следуя модели упомянутого Мак-Артура, происходит эта экологизация за счёт равновесного ущемления собственно биогеографии и превращения её в “селёдку под шубой”. Между тем как экология, так и биогеография, будучи науками многогранными, достаточно сложны для восприятия, даже каждая в отдельности.
Увлечение кладизмом, заимствованным из биологической систематики, привело на некоторое время к возвеличиванию исторической биогеографии в ущерб актуальной. Однако бесконечное многообразие исторических картин распространения таксонов и схем положения материков в прошлом заставляют вновь и вновь возвращаться к современной биогеографической картине, относительно которой только и можно реально оценить многочисленные схемы прошлого и строить прогнозы на будущее.
2.2. Проблема мультитеорийности
Наличие нескольких подходов к одному и тому же предмету исследования нередко приводило науки к кризису. Биогеографии это не грозит, так как она по определению амбивалентна. Четыре теории, обозначенные в предыдущей главе, не исключают, а органически дополняют друг друга, “бомбардируя” единую цель с разных сторон.
Дисперсионная теория (от лат. dispersus – рассеянный, рассыпанный) “заходит на цель” с биологических позиций, отталкиваясь от свойств живых организмов, способствующих расселению. Она мобилизует множество данных о соответствии биологических возможностей организмов тем территориям, где эти возможности наиболее востребованы. Однако структура ареалов, их сравнение и классификация, будучи следствиями причин, заложенных в самих биологических объектах, могут приобрести системный характер лишь после того, как географические свойства живого будут предупредительно сведены в систему. Без этого здание биогеографии можно уподобить архитектурному сооружению, построенному без чертежей с негативными последствиями в критических ситуациях. Поэтому следующая глава имеет цель: придать разнокалиберным данным о географических свойствах жизни системный характер.
Флоро-фаунистическая теория, в отличие от дисперсионной, уходит корнями в физическую географию, которая является ещё одной прародительницей биогеографии. С этой стороны тоже прорабатываются те аспекты, которые другим её направлениям “не по зубам”. Необходимость предварительного ограничения территории для описания, сравнения, типологизации ареалов, анализа флор, фаун, требует системного анализа инфраструктуры географической арены. Простой выход из этого затруднения – использовать какое-нибудь готовое физико-географическое районирование Земли. Однако этот путь, чаще всего, не даёт хороших результатов, ибо организмы в своём распространении руководствуются физико-географическими предпосылками не напрямую – “в лоб”, а опосредованно, через экологические следствия географических причин. Поэтому в последнее время появилось понятие “экография”, т. е. описание и анализ планировки экологических условий на земной поверхности. Однако работы в этом направлении только начались, поэтому ему уделено так много места в этой книге (главы 4, 5, 6, 7). С анализом экографии ареалогия, флористика, фаунистика приобретают системную строгость, а биогеографическое районирование становится объективно универсальным.